The Cerebrum
The iconic gray mantle of the human brain, which appears to make up most of the mass of the brain, is the cerebrum (Figure 1). The wrinkled portion is the cerebral cortex, and the rest of the structure is beneath that outer covering. There is a large separation between the two sides of the cerebrum called the longitudinal fissure. It separates the cerebrum into two distinct halves, a right and left cerebral hemisphere. Deep within the cerebrum, the white matter of the corpus callosum provides the major pathway for communication between the two hemispheres of the cerebral cortex.
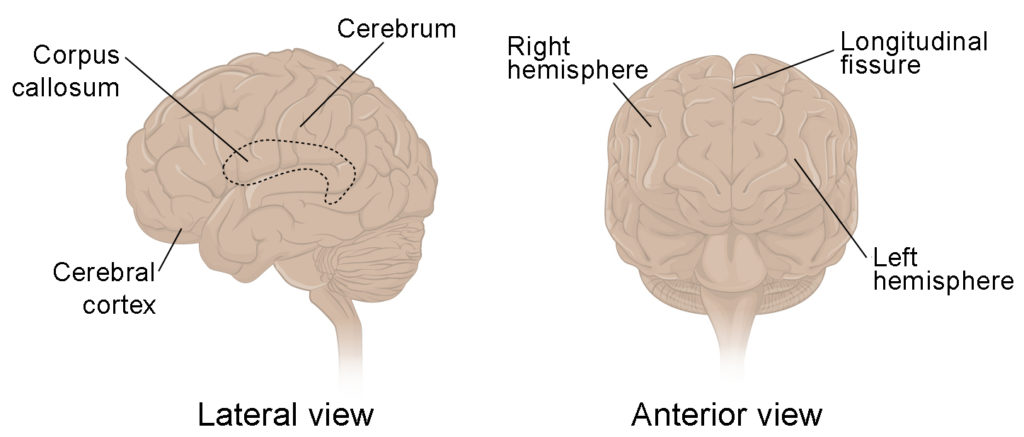
Figure 1. This figure shows the lateral view on the left panel and anterior view on the right panel of the brain. The major parts including the cerebrum are labeled.
Many of the higher neurological functions, such as memory, emotion, and consciousness, are the result of cerebral function. The complexity of the cerebrum is different across vertebrate species. The cerebrum of the most primitive vertebrates is not much more than the connection for the sense of smell. In mammals, the cerebrum comprises the outer gray matter that is the cortex (from the Latin word meaning “bark of a tree”) and several deep nuclei that belong to three important functional groups. The basal nuclei are responsible for cognitive processing, the most important function being that associated with planning movements. The basal forebrain contains nuclei that are important in learning and memory. The limbic cortex is the region of the cerebral cortex that is part of the limbic system, a collection of structures involved in emotion, memory, and behavior.
Cerebral Cortex
The cerebrum is covered by a continuous layer of gray matter that wraps around either side of the forebrain—the cerebral cortex. This thin, extensive region of wrinkled gray matter is responsible for the higher functions of the nervous system. A gyrus (plural = gyri) is the ridge of one of those wrinkles, and a sulcus (plural = sulci) is the groove between two gyri. The pattern of these folds of tissue indicates specific regions of the cerebral cortex. The head is limited by the size of the birth canal, and the brain must fit inside the cranial cavity of the skull. Extensive folding in the cerebral cortex enables more gray matter to fit into this limited space. If the gray matter of the cortex were peeled off of the cerebrum and laid out flat, its surface area would be roughly equal to one square meter. The folding of the cortex maximizes the amount of gray matter in the cranial cavity. During embryonic development, as the telencephalon expands within the skull, the brain goes through a regular course of growth that results in everyone’s brain having a similar pattern of folds. The surface of the brain can be mapped on the basis of the locations of large gyri and sulci. Using these landmarks, the cortex can be separated into four major regions, or lobes (Figure 2).
Lobes of the Cortex
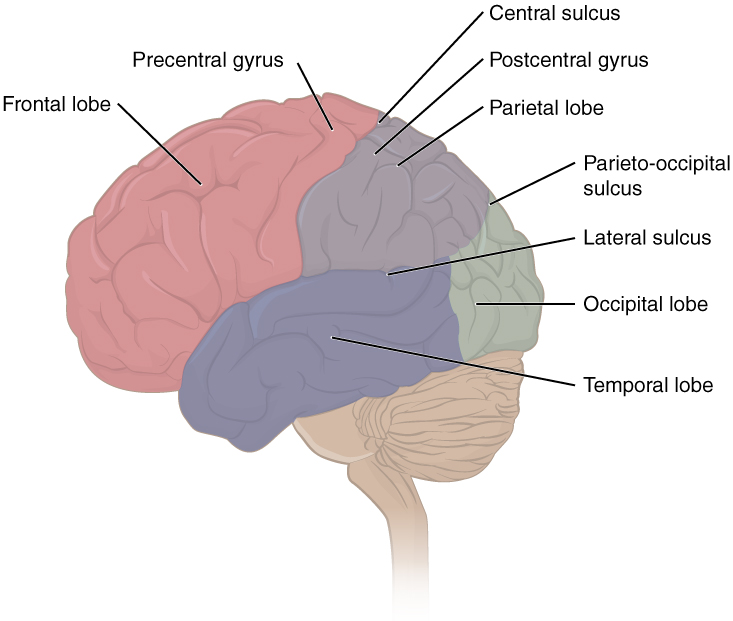
Figure 2. Lobes of the Cerebral Cortex. The cerebral cortex is divided into four lobes. Extensive folding increases the surface area available for cerebral functions.
Areas of the cerebral cortex can be identified as five different lobes. Each lobe plays a role in conscious awareness for one of three functions: Motor areas control voluntary motions, Sensory areas allow for the perception or awareness of different sensations, and Association areas produce meaning from stimuli so that they can be recognized and understood (Figure 3).
The lateral sulcus that separates the temporal lobe from the other regions is one such landmark. Pulling the frontal, temporal lobe, and parietal lobes apart at the lateral sulcus exposes the
– a lobe of cerebral cortex not visible superficially. Superior to the lateral sulcus are the parietal lobe and frontal lobe, which are separated from each other by the central sulcus. The posterior region of the cortex is the occipital lobe, which has no obvious anatomical border between it and the parietal or temporal lobes on the lateral surface of the brain. From the medial surface, an obvious landmark separating the parietal and occipital lobes is called the parieto-occipital sulcus. The fact that there is no obvious anatomical border between these lobes is consistent with the functions of these regions being interrelated.
Insula
The insula possesses a sensory area for taste, and receives signals for sensations from internal organs. Because the insula receives signals from the vestibule of the inner ear, it is also responsible for conscious perception of balance.
Occipital lobe
The occipital lobe is responsible for primary visual perception. It possesses a sensory area for conscious awareness of visual information. It also possesses part of the association area for visual information, so that once you see something you can make sense of the visual information and recognize the object or individual you are looking at. That visual information is complex, so the association area for vision extends into the temporal and parietal lobes as well.
Temporal lobe
The temporal lobe is associated with primary auditory sensation. Because regions of the temporal lobe are part of the limbic system, memory is an important function associated with that lobe. Memory is essentially a sensory function; memories are recalled sensations such as the smell of Mom’s baking or the sound of a barking dog. Even memories of movement are really the memory of sensory feedback from those movements, such as stretching muscles or the movement of the skin around a joint. Structures in the temporal lobe are responsible for establishing long-term memory, but the ultimate location of those memories is usually in the region in which the sensory perception was processed.

Figure 3. Motor, Sensory, and Association areas of the cerebral cortex.
Parietal lobe
The main sensation associated with the parietal lobe is somatosensation, meaning the general sensations associated with the body. Posterior to the central sulcus is the postcentral gyrus, on which is the primary somatosensory cortex. All of the tactile senses are processed in this area, including touch, pressure, tickle, pain, itch, and vibration, as well as more general senses of the body such as proprioception and kinesthesia, which are the senses of body position and movement, respectively. The somatosensory cortex provides an example in which, in essence, the locations of the somatosensory receptors in the body are mapped onto the cerebrum itself. This mapping is often depicted using a sensory homunculus (Figure 4). The term homunculus comes from the Latin word for “little man” and refers to a map of the human body that is laid across a portion of the cerebral cortex.

Figure 4. The Sensory Homunculus A cartoon representation of the sensory homunculus arranged adjacent to the cortical region in which the processing takes place.
In the somatosensory cortex, the external genitals, feet, and lower legs are represented on the medial face of the gyrus within the longitudinal fissure. As the gyrus curves out of the fissure and along the surface of the parietal lobe, the body map continues through the thighs, hips, trunk, shoulders, arms, and hands. The head and face are just lateral to the fingers as the gyrus approaches the lateral sulcus.
The representation of the body in this topographical map is medial to lateral from the lower to upper body. It is a continuation of the topographical arrangement seen in the dorsal column system, where axons from the lower body are carried in the fasciculus gracilis, whereas axons from the upper body are carried in the fasciculus cuneatus. As the dorsal column system continues into the medial lemniscus, these relationships are maintained. Also, the head and neck axons running from the trigeminal nuclei to the thalamus run adjacent to the upper body fibers. The connections through the thalamus maintain topography such that the anatomic information is preserved.
Note that this correspondence does not result in a perfectly miniature scale version of the body, but rather exaggerates the more sensitive areas of the body, such as the fingers and lower face. Less sensitive areas of the body, such as the shoulders and back, are mapped to smaller areas on the cortex.
Frontal lobe
Anterior to this region are a few areas that are associated with planned movements. The premotor area is responsible for thinking of a movement to be made. The frontal eye fields are important in eliciting eye movements and in attending to visual stimuli. Broca’s area is responsible for the production of language, or controlling movements responsible for speech; in the vast majority of people, it is located only on the left side. Anterior to these regions is the prefrontal lobe, which serves cognitive functions that can be the basis of personality, short-term memory, and consciousness. The prefrontal lobotomy is an outdated mode of treatment for personality disorders (psychiatric conditions) that profoundly affected the personality of the patient.
The completion of cortical processing through the primary, associative, and integrative sensory areas initiates a similar progression of motor processing, usually in different cortical areas. While the sensory cortical areas are located in the insula, occipital, temporal, and parietal lobes, motor functions are largely controlled by the frontal lobe.
The most anterior regions of the frontal lobe—the prefrontal areas—are important for executive functions, which are those cognitive functions that lead to goal-directed behaviors. These higher cognitive processes include working memory, which has been called a “mental scratch pad,” that can help organize and represent information that is not in the immediate environment.

Figure 5. Phineas Gage. The victim of an accident while working on a railroad in 1848, Phineas Gage had a large iron rod impaled through the prefrontal cortex of his frontal lobe. After the accident, his personality appeared to change, but he eventually learned to cope with the trauma and lived as a coach driver even after such a traumatic event. (credit b: John M. Harlow, MD)
The prefrontal lobe is responsible for aspects of attention, such as inhibiting distracting thoughts and actions so that a person can focus on a goal and direct behavior toward achieving that goal. The functions of the prefrontal cortex are integral to the personality of an individual, because it is largely responsible for what a person intends to do and how they accomplish those plans.
A famous case of damage to the prefrontal cortex is that of Phineas Gage, dating back to 1848. He was a railroad worker who had a metal spike impale his prefrontal cortex (Figure 5). He survived the accident, but according to second-hand accounts, his personality changed drastically.
Friends described him as no longer acting like himself. Whereas he was a hardworking, amiable man before the accident, he turned into an irritable, temperamental, and lazy man after the accident. Many of the accounts of his change may have been inflated in the retelling, and some behavior was likely attributable to alcohol used as a pain medication. However, the accounts suggest that some aspects of his personality did change. There is new evidence that though his life changed dramatically, he was able to become a functioning stagecoach driver, suggesting that the brain has the ability to recover even from major trauma such as this.
Secondary Motor Cortices
In generating motor responses, the executive functions of the prefrontal cortex will need to initiate actual movements. One way to define the prefrontal area is any region of the frontal lobe that does not elicit movement when electrically stimulated. These are primarily in the anterior part of the frontal lobe. The regions of the frontal lobe that remain are the regions of the cortex that produce movement.
The prefrontal areas project into the secondary motor cortices, which include the premotor cortex and the supplemental motor area. Two important regions that assist in planning and coordinating movements are located adjacent to the primary motor cortex. The premotor cortex is more lateral, whereas the supplemental motor area is more medial and superior. The premotor area aids in controlling movements of the core muscles to maintain posture during movement, whereas the supplemental motor area is hypothesized to be responsible for planning and coordinating movement. The supplemental motor area also manages sequential movements that are based on prior experience (that is, learned movements). Neurons in these areas are most active leading up to the initiation of movement.
For example, these areas might prepare the body for the movements necessary to drive a car in anticipation of a traffic light changing. Adjacent to these two regions are two specialized motor planning centers. The frontal eye fields are responsible for moving the eyes in response to visual stimuli. There are direct connections between the frontal eye fields and the superior colliculus. Also, anterior to the premotor cortex and primary motor cortex is Broca’s area. This area is responsible for controlling movements of the structures of speech production. The area is named after a French surgeon and anatomist who studied patients who could not produce speech. They did not have impairments to understanding speech, only to producing speech sounds, suggesting a damaged or underdeveloped Broca’s area.
Primary Motor Cortex
The primary motor cortex is located in the precentral gyrus of the frontal lobe. Walter Penfield, a neurosurgeon, described much of the basic understanding of the primary motor cortex by electrically stimulating the surface of the cerebrum. Penfield would probe the surface of the cortex while the patient was only under local anesthesia so that he could observe responses to the stimulation. This led to the belief that the precentral gyrus directly stimulated muscle movement. We now know that the primary motor cortex receives input from several areas that aid in planning movement, and its principle output stimulates spinal cord neurons to stimulate skeletal muscle contraction.
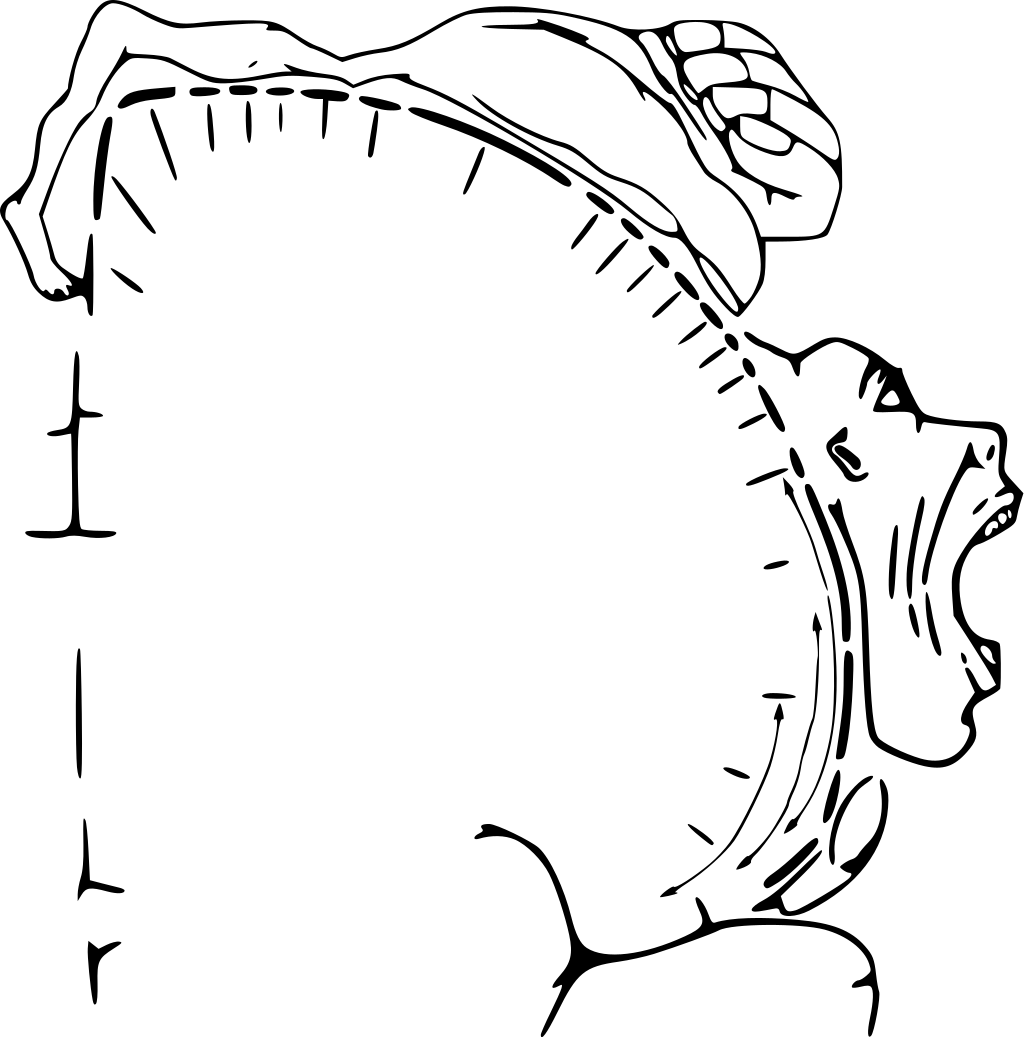
Figure 6. Motor homunculus over the precentral gyrus of the frontal lobe, shown cut in a frontal section.
The primary motor cortex is arranged in a similar fashion to the primary somatosensory cortex, in that it has a topographical map of the body, creating a motor homunculus (Figure 6). The neurons responsible for musculature in the feet and lower legs are in the medial wall of the precentral gyrus, with the thighs, trunk, and shoulder at the crest of the longitudinal fissure. The hand and face are in the lateral face of the gyrus.
Additionally, the relative space allotted for the different regions is exaggerated in muscles that have greater enervation. The greatest amount of cortical space is given to muscles that perform fine, agile movements, such as the muscles of the fingers and the lower face. The “power muscles” that perform coarser movements, such as the buttock and back muscles, occupy much less space on the motor cortex.
White matter of the Cerebral Cortex
Grey matter of the cerebral cortex constantly communicates with other areas of the cotrex, as well as with inferior areas of the brain. To do this, three different types of tracts carry signals from axons (fibers) to different destinations (Figure 7). Association fibers link areas of grey matter in the same cerebral hemisphere. Commissural fibers link areas of grey matter in opposite cerebral hemispheres, the corpus callosum is the largest area of commissural fibers. The cererbrum communicates with deeper areas of grey matter like the basal nuclei, or with inferior areas of the brain, by way of projection fibers.

Figure 7. Projection, Association, and Commissural fibers.
Subcortical Structures

Figure 8. Frontal Section of Cerebral Cortex and Basal Nuclei. The major components of the basal nuclei, shown in a frontal section of the brain, are the caudate (just lateral to the lateral ventricle), the putamen (inferior to the caudate and separated by the large white-matter structure called the internal capsule), and the globus pallidus (medial to the putamen).
Beneath the cerebral cortex are sets of nuclei known as subcortical nuclei that augment cortical processes. The nuclei of the basal forebrain serve as the primary location for acetylcholine production, which modulates the overall activity of the cortex, possibly leading to greater attention to sensory stimuli. Alzheimer’s disease is associated with a loss of neurons in the basal forebrain.
Basal Nuclei
The basal nuclei are a set of nuclei in the cerebrum responsible for comparing cortical processing with the general state of activity in the nervous system to influence the likelihood of movement taking place. For example, while a student is sitting in a classroom listening to a lecture, the basal nuclei will keep the urge to jump up and scream from actually happening. (The basal nuclei are also referred to as the basal ganglia, although that is potentially confusing because the term ganglia is typically used for peripheral structures.)
The major structures of the basal nuclei that control movement are the caudate, putamen, and globus pallidus, which are located deep in the cerebrum. The caudate is a long nucleus that follows the basic C-shape of the cerebrum from the frontal lobe, through the parietal and occipital lobes, into the temporal lobe. The putamen is mostly deep in the anterior regions of the frontal and parietal lobes. Together, the caudate and putamen are called the striatum. The globus pallidus is a layered nucleus that lies just medial to the putamen; they are called the lenticular nuclei because they look like curved pieces fitting together like lenses. The globus pallidus has two subdivisions, the external and internal segments, which are lateral and medial, respectively. These nuclei are depicted in a frontal section of the brain in Figure 8.

Figure 9. Connections of Basal Nuclei. Input to the basal nuclei is from the cerebral cortex, which is an excitatory connection releasing glutamate as a neurotransmitter. This input is to the striatum, or the caudate and putamen. In the direct pathway, the striatum projects to the internal segment of the globus pallidus and the substantia nigra pars reticulata (GPi/SNr). This is an inhibitory pathway, in which GABA is released at the synapse, and the target cells are hyperpolarized and less likely to fire. The output from the basal nuclei is to the thalamus, which is an inhibitory projection using GABA.
The basal nuclei in the cerebrum are connected with a few more nuclei in the brain stem that together act as a functional group that forms a motor pathway. Two streams of information processing take place in the basal nuclei. All input to the basal nuclei is from the cortex into the striatum (Figure 9).
The direct pathway is the projection of axons from the striatum to the globus pallidus internal segment (GPi) and the substantia nigra pars reticulata (SNr). The GPi/SNr then projects to the thalamus, which projects back to the cortex. The indirect pathway is the projection of axons from the striatum to the globus pallidus external segment (GPe), then to the subthalamic nucleus (STN), and finally to GPi/SNr. The two streams both target the GPi/SNr, but one has a direct projection and the other goes through a few intervening nuclei. The direct pathway causes the disinhibition of the thalamus (inhibition of one cell on a target cell that then inhibits the first cell), whereas the indirect pathway causes, or reinforces, the normal inhibition of the thalamus. The thalamus then can either excite the cortex (as a result of the direct pathway) or fail to excite the cortex (as a result of the indirect pathway).
The switch between the two pathways is the substantia nigra pars compacta, which projects to the striatum and releases the neurotransmitter dopamine. Dopamine receptors are either excitatory (D1-type receptors) or inhibitory (D2-type receptors). The direct pathway is activated by dopamine, and the indirect pathway is inhibited by dopamine.
When the substantia nigra pars compacta is firing, it signals to the basal nuclei that the body is in an active state, and movement will be more likely. When the substantia nigra pars compacta is silent, the body is in a passive state, and movement is inhibited. To illustrate this situation, while a student is sitting listening to a lecture, the substantia nigra pars compacta would be silent and the student less likely to get up and walk around. Likewise, while the professor is lecturing, and walking around at the front of the classroom, the professor’s substantia nigra pars compacta would be active, in keeping with his or her activity level.
Watch this video to learn about the basal nuclei (also known as the basal ganglia), which have two pathways that process information within the cerebrum.
As shown in this video, the direct pathway is the shorter pathway through the system that results in increased activity in the cerebral cortex and increased motor activity. The direct pathway is described as resulting in “disinhibition” of the thalamus. What does disinhibition mean? What are the two neurons doing individually to cause this?
Disorders of the Basal Nuclei
Parkinson’s disease is a disorder of the basal nuclei, specifically of the substantia nigra, that demonstrates the effects of the direct and indirect pathways. Parkinson’s disease is the result of neurons in the substantia nigra pars compacta dying. These neurons release dopamine into the striatum. Without that modulatory influence, the basal nuclei are stuck in the indirect pathway, without the direct pathway being activated. The direct pathway is responsible for increasing cortical movement commands. The increased activity of the indirect pathway results in the hypokinetic disorder of Parkinson’s disease. Parkinson’s disease is neurodegenerative, meaning that neurons die that cannot be replaced, so there is no cure for the disorder. Treatments for Parkinson’s disease are aimed at increasing dopamine levels in the striatum. Currently, the most common way of doing that is by providing the amino acid L-DOPA, which is a precursor to the neurotransmitter dopamine and can cross the blood-brain barrier. With levels of the precursor elevated, the remaining cells of the substantia nigra pars compacta can make more neurotransmitter and have a greater effect. Unfortunately, the patient will become less responsive to L-DOPA treatment as time progresses, and it can cause increased dopamine levels elsewhere in the brain, which are associated with psychosis or schizophrenia.
Visit this site for a thorough explanation of Parkinson’s disease.
Compared with the nearest evolutionary relative, the chimpanzee, the human has a brain that is huge. At a point in the past, a common ancestor gave rise to the two species of humans and chimpanzees. That evolutionary history is long and is still an area of intense study. But something happened to increase the size of the human brain relative to the chimpanzee. Read this article in which the author explores the current understanding of why this happened. According to one hypothesis about the expansion of brain size, what tissue might have been sacrificed so energy was available to grow our larger brain? Based on what you know about that tissue and nervous tissue, why would there be a trade-off between them in terms of energy use?
Limbic System
The limbic system refers to an area of deep grey and white matter structures that, while anatomically near one anot

Figure 10. Structures of the limbic system and nearby diencephalon.
her are not necessarily linked in shared function (Figure 10). While the term limbic system is somewhat antiquated for this reason, it remains useful to refer to this group of structures as a region of the brain. The hippocampus is one of the few areas of the brain in which continuous mitosis of neurons occurs. Because new neurons are constantly produced in this area, the hippocampus is used to generate new memories, in
conjunction with the hypothalamus. The hiipocampus also is one of the first areas of the cerebrum to receive olfactory information from the olfactory tract. As a result, the hippocampus generates a close link between smells and specific memories. The amygdala is a structure of the limbic system that becomes activated when someone is surprised by something, or when something catches one’s attention. Long thought to be the seat of fear responses, the amgydala is now understood to have a more generalized function. The cingulate cortex (or cingulate gyrus) sits just superficial to the corpus callosum. Recent research suggests that the cingulate cortex processes emotional responses, helps to link those emotions to endocrine responses in the body, reward-based decision making, imagination, and memory. The fornix is an area of grey matter in the limbic system that links it to the structures of the diencephalon.
Everyday Connections: The Myth of Left Brain/Right Brain
There is a persistent myth that people are “right-brained” or “left-brained,” which is an oversimplification of an important concept about the cerebral hemispheres. There is some lateralization of function, in which the left side of the brain is devoted to language function and the right side is devoted to spatial and nonverbal reasoning. Whereas these functions are predominantly associated with those sides of the brain, there is no monopoly by either side on these functions. Many pervasive functions, such as language, are distributed globally around the cerebrum. Some of the support for this misconception has come from studies of split brains.
A drastic way to deal with a rare and devastating neurological condition (intractable epilepsy) is to separate the two hemispheres of the brain. After sectioning the corpus callosum, a split-brained patient will have trouble producing verbal responses on the basis of sensory information processed on the right side of the cerebrum, leading to the idea that the left side is responsible for language function. However, there are well-documented cases of language functions lost from damage to the right side of the brain.
The deficits seen in damage to the left side of the brain are classified as aphasia, a loss of speech function; damage on the right side can affect the use of language. Right-side damage can result in a loss of ability to understand figurative aspects of speech, such as jokes, irony, or metaphors. Nonverbal aspects of speech can be affected by damage to the right side, such as facial expression or body language, and right-side damage can lead to a “flat affect” in speech, or a loss of emotional expression in speech—sounding like a robot when talking.
Candela Citations
- Motor and Sensory Regions of the Cerebral Cortex.. Authored by: Blausen.com staff (2014). . Provided by: Medical gallery of Blausen Medical 2014. WikiJournal of Medicine . Located at: https://commons.wikimedia.org/wiki/File:Blausen_0102_Brain_Motor%26Sensory.png. License: CC BY: Attribution
- Anatomy & Physiology. Authored by: OpenStax College. Provided by: Rice University. Located at: http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@9.1. License: CC BY: Attribution. License Terms: Download for free at http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@9.1
- Coronal T2 (grey scale inverted) MRI of the brain at the level of the the caudate nuclei emphasizing corpus callosum.. Authored by: Frank Gaillard. Located at: https://commons.wikimedia.org/wiki/File:26638.medium-emphasizing-corpus-callosum.png. License: CC BY-SA: Attribution-ShareAlike
- Limbic System. . Authored by: Blausen.com staff (2014). . Provided by: Medical gallery of Blausen Medical 2014. WikiJournal of Medicine . Located at: https://commons.wikimedia.org/wiki/File:Blausen_0614_LimbicSystem.png. License: CC BY: Attribution