Smooth muscle (so-named because the cells do not have striations) is present in the walls of hollow organs like the urinary bladder, uterus, stomach, intestines, and in the walls of passageways, such as the arteries and veins of the circulatory system, and the tracts of the respiratory, urinary, and reproductive systems (Figure 1). Smooth muscle is also present in the eyes, where it functions to change the size of the iris and alter the shape of the lens; and in the skin where it causes hair to stand erect in response to cold temperature or fear.

Figure 1. Smooth Muscle Tissue. Smooth muscle tissue is found around organs in the digestive, respiratory, reproductive tracts and the iris of the eye. LM × 1600. (Micrograph provided by the Regents of University of Michigan Medical School © 2012)
Smooth muscle fibers are spindle-shaped (wide in the middle and tapered at both ends, somewhat like a football) and have a single nucleus; they range from about 30 to 200 μm (thousands of times shorter than skeletal muscle fibers), and they produce their own connective tissue, endomysium. Although they do not have striations and sarcomeres, smooth muscle fibers do have actin and myosin contractile proteins, and thick and thin filaments. These thin filaments are anchored by dense bodies. A dense body is analogous to the Z-discs of skeletal and cardiac muscle fibers and is fastened to the sarcolemma. When the thin filaments slide past the thick filaments, they pull on the dense bodies, structures tethered to the sarcolemma, which then pull on the intermediate filaments networks throughout the sarcoplasm. This arrangement causes the entire muscle fiber to contract in a manner whereby the ends are pulled toward the center, causing the midsection to bulge in a corkscrew motion (Figure 2).
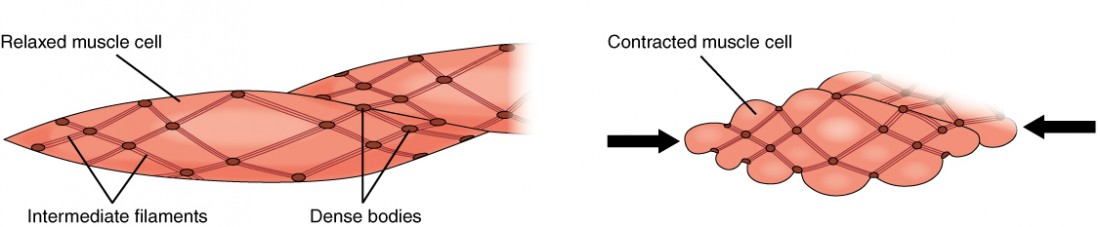
Figure 2. Muscle Contraction. The dense bodies and intermediate filaments are networked through the sarcoplasm, which cause the muscle fiber to contract.

Figure 3. Motor Units. A series of axon-like swelling, called varicosities or “boutons,” from autonomic neurons form motor units through the smooth muscle.
Smooth muscle is organized in two ways: as single-unit smooth muscle, which is much more common; and as multiunit smooth muscle. The two types have different locations in the body and have different characteristics.
Single-unit muscle has its muscle fibers joined by gap junctions so that the muscle contracts as a single unit. This type of smooth muscle is found in the walls of all visceral organs except the heart (which has cardiac muscle in its walls), and so it is commonly called visceral muscle. Because the muscle fibers are not constrained by the organization and stretchability limits of sarcomeres, visceral smooth muscle has a stress-relaxation response. This means that as the muscle of a hollow organ is stretched when it fills, the mechanical stress of the stretching will trigger contraction, but this is immediately followed by relaxation so that the organ does not empty its contents prematurely. This is important for hollow organs, such as the stomach or urinary bladder, which continuously expand as they fill. The smooth muscle around these organs also can maintain a muscle tone when the organ empties and shrinks, a feature that prevents “flabbiness” in the empty organ. In general, visceral smooth muscle produces slow, steady contractions that allow substances, such as food in the digestive tract, to move through the body.
Multiunit smooth muscle cells rarely possess gap junctions, and thus are not electrically coupled. As a result, contraction does not spread from one cell to the next, but is instead confined to the cell that was originally stimulated. Stimuli for multiunit smooth muscles come from autonomic nerves or hormones but not from stretching. This type of tissue is found around large blood vessels, in the respiratory airways, and in the eyes.
Hyperplasia in Smooth Muscle
Similar to skeletal and cardiac muscle cells, smooth muscle can undergo hypertrophy to increase in size. Unlike other muscle, smooth muscle can also divide to produce more cells, a process called hyperplasia. This can most evidently be observed in the uterus at puberty, which responds to increased estrogen levels by producing more uterine smooth muscle fibers, and greatly increases the size of the myometrium.
Cardiac muscle tissue
Cardiac muscle tissue is only found in the heart. Highly coordinated contractions of cardiac muscle pump blood into the vessels of the circulatory system. Similar to skeletal muscle, cardiac muscle is striated and organized into sarcomeres, possessing the same banding organization as skeletal muscle.
However, cardiac muscle fibers are shorter than skeletal muscle fibers and usually contain only one nucleus, which is located in the central region of the cell. Cardiac muscle fibers also possess many mitochondria and myoglobin, as ATP is produced primarily through aerobic metabolism. Cardiac muscle fibers cells also are extensively branched and are connected to one another at their ends by intercalated discs. An intercalated disc allows the cardiac muscle cells to contract in a wave-like pattern so that the heart can work as a pump.
Intercalated discs are part of the sarcolemma and contain two structures important in cardiac muscle contraction: gap junctions and desmosomes. A gap junction forms channels between adjacent cardiac muscle fibers that allow the depolarizing current produced by cations to flow from one cardiac muscle cell to the next. This joining is called electric coupling, and in cardiac muscle it allows the quick transmission of action potentials and the coordinated contraction of the entire heart. This network of electrically connected cardiac muscle cells creates a functional unit of contraction called a syncytium. The remainder of the intercalated disc is composed of desmosomes. A desmosome is a cell structure that anchors the ends of cardiac muscle fibers together so the cells do not pull apart during the stress of individual fibers contracting (Figure 4).

Figure 4. Cardiac Muscle. Intercalated discs are part of the cardiac muscle sarcolemma and they contain gap junctions and desmosomes.
Contractions of the heart (heartbeats) are controlled by specialized cardiac muscle cells called pacemaker cells that directly control heart rate. Although cardiac muscle cannot be consciously controlled, the pacemaker cells respond to signals from the autonomic nervous system (ANS) to speed up or slow down the heart rate. The pacemaker cells can also respond to various hormones that modulate heart rate to control blood pressure.
The wave of contraction that allows the heart to work as a unit, called a functional syncytium, begins with the pacemaker cells. This group of cells is self-excitable and able to depolarize to threshold and fire action potentials on their own, a feature called autorhythmicity; they do this at set intervals which determine heart rate. Because they are connected with gap junctions to surrounding muscle fibers and the specialized fibers of the heart’s conduction system, the pacemaker cells are able to transfer the depolarization to the other cardiac muscle fibers in a manner that allows the heart to contract in a coordinated manner.
Candela Citations
- Chapter 10. Authored by: OpenStax College. Provided by: Rice University. Located at: http://cnx.org/contents/14fb4ad7-39a1-4eee-ab6e-3ef2482e3e22@15.1.. Project: Anatomy & Physiology. License: CC BY: Attribution. License Terms: Download for free at http://cnx.org/content/col11496/latest/.