Homo heidelbergensis (<600 kya)
(“same” / Heidelberg, Germany)

Figure 33.1 Model of Homo heidelbergensis head and shoulders. “Homo heidelbergensis adult male – head model – Smithsonian Museum of Natural History – 2012-05-17” by Tim Evanson is licensed under CC BY-SA 2.0<.
SITES
Europe:
Spain: Sima de los Huesos
France: Arago
Germany: Mauer, Steinheim, Ehringsdorf, Schöningen, Bilzingsleben
England: Boxgrove, Swanscombe
Italy: Ciampate del Diavolo (footprints)
Hungary: Vértesszöllös
Greece: Petralona, Apidima
Africa:
Morocco: Jebel Irhoud
Ethiopia: Bodo, Omo (Kibish Beds)
Zambia: Kabwe
Tanzania: Lake Eyasi, Laetoli (Ngaloba Beds)
South Africa: Florisbad, Elandsfontein
Asia:
China: Dali, Maba, Jinniushan
PEOPLE
Otto Schoetensack and many more
INTRODUCTION
For many years, fossil material from ~500–200 kya from Africa, Asia, and Europe that was more human- or sapiens-like was included in our own genus and species but was distinguished as “Early Archaic” Homo sapiens (EAHS). There was much debate as to when to draw the line between more erectus-like forms and more sapiens-like forms. The prevailing view was that material on all three continents was descended from H. erectus. (The various geographic species distinctions for H. erectus had not yet come into use.) Again, this is referred to as the Regional Continuity Model (RCM) for the origin of modern humans. In this scenario, erectus-like forms on each of the continents slowly evolved into modern humans via gene flow between populations. This is in contrast to the Recent African Origin (RAO) model whereby modern humans evolved in Africa and moved out to eventually replace archaic forms elsewhere, e.g. H. erectus in Asia.
The RAO model has gained in popularity due to a combination of the reevaluation of fossil material and especially DNA methods aimed at evaluating genetic distance between species, in terms of number of years since divergence from a common ancestor. The material from Asia that had previously been assigned to EAHS has been relegated to H. erectus, with very little evidence for an intermediate form bridging the gap between H. erectus and AMH.
Fossil material from Europe and Africa that was formerly assigned to EAHS is now termed Homo heidelbergensis, with some researchers using Homo rhodesiensis for some of the African material. It is now well accepted that H. heidelbergensis was ancestral to both humans and neandertals (see Figure 33.2—note that in this scenario, H. antecessor is not considered to be ancestral to H. heidelbergensis).

Figure 33.2 Human phylogeny. “Homo-Stammbaum, Version Stringer” by Chris Stringer is licensed under CC BY-SA 3.0 DE.
PHYLOGENY
H. heidelbergensis is thought to have evolved from a more derived form of H. ergaster. Some researchers believe that the material from Tighenif (formerly Ternifine), Algeria, known by some as Homo mauritanicus, is a possible transitional form between the two species. Mandibles from Tighenif are very similar to the type specimen, the Mauer mandible (see Figure 33.3) from the Heidelberg area of Germany, from which the species name is derived. In addition to the material from North Africa, the oldest material is from the Bodo site in Ethiopia, dated to 600 kya. Thus while an African origin is favored, some believe that H. heidelbergensis is descended from H. antecessor in Europe. Whether H. heidelbergensis evolved in Europe or Africa, they had to have migrated from one continent to the other. Thus if H. heidelbergensis evolved in Europe from H. antecessor, a portion of the population migrated to Africa and evolved toward AMH, leaving some regional population of the remaining European stock to evolve into H. neanderthalensis. The other more popular scenario has H. heidelbergensis evolving in Africa from some descendant form of Homo ergaster. A portion of the population then emigrated to Europe and evolved into neandertals while a portion of the remaining African stock evolved toward AMH.
In addition, a new species of hominin is also thought to be descended from H. heidelbergensis. The Denisovans (see this chapter), as they have come to be known due to their discovery in the Denisova Cave in the Altai Mountains of Russia, are thought to have branched off from the H. heidelbergensis lineage that led to neandertals. DNA analyses show that Denisovans interbred with neandertals, as well as the first wave of AMH that left Africa, possibly around 125 kya and subsequently settled Melanesia and Australia.

Figure 33.3 Type specimen: Mauer mandible. “Unterkiefer von Mauer (Replika)” by Gerbil is licensed under CC BY-SA 3.0.
Contrary new DNA evidence suggests that H. heidelbergensis branched off from some common ancestors prior to 800 kya and is thus not part of our recent lineage. H. heidelbergensis then divided into the neandertals and the Denisovan lineages, but interbreeding continued between them in areas of geographic overlap.
There is good evidence that H. heidelbergensis gave rise to neandertals. There are skulls from the sites of Erhingsdorf, Germany; Fontechavade, France; and Sima de los Huesos, Spain, that appear to be transitional in that they exhibit characteristics seen in neandertals, such as similar brow ridge morphology, enlarged facial sinuses, occipital bun, etc. It is rare in paleoanthropology to have such good support for continuity in physical characteristics between ancestor and descendent species. However, the fact that those species lived more recently in time and in developed and populated areas certainly helps, in terms of facilitating discoveries.

Figure 33.4 Homo heidelbergensis from Steinheim, Germany. “Homo steinheimensis, holotype” by Dr. Günter Bechly is licensed under CC BY-SA 3.0.
DISCOVERY AND GEOGRAPHIC RANGE
The earliest discoveries of H. heidelbergensis are from Germany. The type specimen was discovered in 1907 in Mauer, Germany. The oldest site is Bodo, Ethiopia (600 kya). There are numerous H. heidelbergensis sites in Europe (e.g. Steinheim, see Figure 33.4) that date from as early as 500 kya and range from Spain through Eastern Europe. The greatest number of individuals came from the Sima de los Huesos (“Pit of Bones”) (see Figure 33.5) site in the Atapuerca Mountains of Spain. Both H. antecessor and H. heidelbergensis sites in that area were discovered when a railway was built. The H. heidelbergensis remains were found in a deep chamber inside of a cave, hence the name, “Pit of Bones.” More than 32 individuals have been recovered, and most of them are juveniles. While there has been much speculation, the significance of this find in terms of cultural complexity (see ENVIRONMENT AND WAY OF LIFE below) remains elusive. There are a couple of sites in Asia that some researchers believe are also representative of H. heidelbergensis, including Dali, China. Sites in Africa (also referred to as H. rhodesiensis) include Jebel Irhoud in North Africa; Omo, Bodo, Ndutu, Eyasi, and Ngaloba in East Africa; Kabwe in Zambia; and Florisbad and Elandsfontein in South Africa.

Figure 33.5 Homo heidelbergensis from Sima de los Huesos, Spain. “Homo heidelbergensis-Cranium -5” by José-Manuel Benito Álvarez is licensed under CC BY-SA 2.5.
PHYSICAL CHARACTERISTICS
In general, H. heidelbergensis was less robust, both cranially and postcranially, than the erectus grade and more robust than AMH. There is intraspecific variability in craniofacial morphology, with some specimens being more robust and others more gracile. While this is not surprising, considering the temporal and geographic range for the species, it has led to disagreement as to how to define it.
H. heidelbergensis is primarily distinguished from erectus-like forms by its increased cranial capacity (1100–1400 cc—93% that of AMH) and more modern skull vault. Cerebral expansion, especially of the parietal lobes, led to increased cranial breadth in the superior aspect of the skull vault and thus a more vertically oriented skull. The occipital region is less angular due to reduced robusticity in the nuchal musculature. Some specimens have very pronounced brow ridges (see Figure 33.7) and some have speculated that those individuals represent males of the species. They share the presence of a canine fossa with AMH. The canine fossa is a depression above the canine on either side of the maxilla. The robusticity of their jaw morphology and tooth wear suggests that they were using their mouths and teeth as tools, for example, pulling meat off bones and working hides. Surprisingly, the middle ear bones were discovered for one specimen, and it could thus be determined that their hearing was comparable with our own (Martínez et al. 2004).

Figure 33.6 Homo heidelbergensis by Keenan Taylor.
Some H. heidelbergensis are considered to be transitional and exhibited characteristics seen in the subsequent neandertals. Their midfacial region was (1) prognathic with a corresponding large nose and (2) puffy due to large frontal and maxillary sinuses. Their zygomatics have a “swept back” appearance, and the posterior skull vault exhibited an occipital bun. The outer layer of their long bones (the cortical bone) was thick, and the interior medullary (marrow) cavity was narrow. The species was taller than neandertals, with males and females averaging 5′ 9″ (175 cm) and 5′ 2″ (157 cm), respectively. Average weights for males and females were 136 lb (62 kg) and 112 lb (51 kg), respectively (Smithsonian Institution 2015).
Review of Derived Characteristics
- Increase in brain size & complexity of skull architecture, and corresponding vault changes.
- Loss of angulation in occipital region.
- Canine fossa.
Review of Characteristics Inherited by Neandertals
- Midfacial prognathism and large nose.
- Puffy appearance to face due to large frontal and maxillary sinuses.
- “Swept back” zygomatics.
- Occipital bun.
- Thick cortical bone and narrow medullary cavity.

Figure 33.7“Broken Hill Man” from Kabwe, Zambia (formerly “Rhodesian Man”). “Broken Hill Skull (Replica01)” by Gerbil is licensed under CC BY-SA 3.0.
ENVIRONMENT AND WAY OF LIFE
The Bodo site was of course, close to the equator and hence warm. H. heidelbergensis moved into Europe during the Holstein interglacial period (400–300 kya) (Wikipedia contributors 2015e).

Figure 33.8 Homo heidelbergensis hunting by Keenan Taylor.
Like those species that preceded them, H. heidelbergensis were mobile foragers. They left evidence for both seasonal and differential use camps. In addition to using rock shelters and caves for shelter, they are the first species for which we have evidence of building free-standing structures. At the site of Terra Amata in the south of France, the living floors of free-standing structures have been excavated. It is thought that a group returned to the site annually for fishing and other subsistence activities and reconstructed their huts (up to 11 times) on the exact same site (see Figure 33.9). They are credited as having been the first to make the tools necessary for efficient fishing. Like past species, their big-game hunting capabilities are questionable. However, there is evidence that they may have ambushed large animals by forcing them off cliffs or cornering them in dead-end canyons. Support for ambush comes from faunal assemblages on the Channel Islands off the coast of France. The remains are from animals in prime condition, and the frequency of the various bones shows that the hominins were differentially removing the limbs and bringing them back to butcher at a home base. Javelin-type spears and thousands of faunal remains have been recovered at Schöningen, Germany. The spears were fire-hardened for strength and expertly weighted for flight, like modern javelins.

Figure 33.9 Hut design at Terra Amata. “Terra-Amata-Hut” by Locutus Borg is in the public domain.
While some tools from H. heidelbergensis are Oldowan-like, most are of the Acheulian tradition. The species is credited with inventing a more conservative method, termed the Levallois technique, for controlling flake shape and maximizing their yield from a core. Flakes could then be worked into a variety of tools. They could also shape the core in such a way that a point could be struck off that was sharp on all sides (see Figure 33.10). H. heidelbergensis were the first to make compound tools, i.e. tools with more than one component, such as hammers and stone-tipped spears. There is a great website demonstrating and describing tool production at http://donsmaps.com/makingflinttools.html, as well as an animated visual for the Levallois technique at http://en.wikipedia.org/wiki/Levallois_technique.
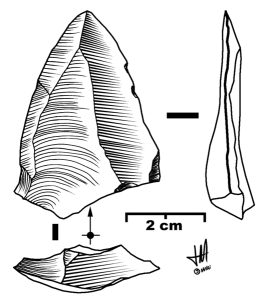
Figure 33.10 Levallois point. “Levallois point” by José-Manuel Benito Álvarez is licensed under CC BY-SA 2.5.
H. heidelbergensis is the first species for which there is ample evidence of the controlled use of fire, in that hearths have been found at several sites. In addition to the aforementioned inventions, a couple of novel cultural practices have been suggested for the species. They may have made and used furniture, such as seaweed beds and stone blocks, and there is some evidence of art or written communication in the form of arcs and angles and the use of ocher (mineral pigments). A fine pink quartz hand axe, nicknamed “Excalibur,” was found among the bodies in the “Pit of Bones.” Some researchers believe it is the earliest evidence of ritual associated with burial, in that the artifact was seemingly unused and manufactured from exotic stone. The seclusion of the bodies may also represent an attempt at keeping them from being ravaged by scavengers. All of these advancements and innovations are unequivocal support for the increase in cognition that resulted from the degree of encephalization and changes in brain architecture that are evident in the skull size and shape of H. heidelbergensis. Finally, if the new DNA evidence is correct and H. heidelbergensis branched off from our ancestry versus being our ancestor, the behavioral and cultural complexity apparent at H. heidelbergensis sites indicates that our common ancestor was cognitively advanced more than 800 kya!

Figure 33.11 Hand axe from Boxgrove, England. “Boxgrove handaxe” by Midnightblueowl is licensed under CC BY-SA 3.0.
Candela Citations
- The History of our Tribe: Hominini. Authored by: Barbara Welker. Provided by: SUNY Geneseo. Located at: https://milnepublishing.geneseo.edu/the-history-of-our-tribe-hominini/. Project: Open SUNY Textbooks. License: CC BY-NC-SA: Attribution-NonCommercial-ShareAlike

{kind=link}
{kind=link}
.JPG){kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}