Describe and explain a range of mechanisms for generating genetic diversity
Now that we know how meiosis works, let’s see how it and its involved processes contribute to genetic diversity.
Learning Objectives
- Understand how sexual reproduction leads to different sexual life cycles
- Understand how meiosis contributes to genetic diversity
- Understand the impact of translocation between non-homologous chromosomes during meiosis or mitosis
- Understand how mitosis, meiosis, and random fertilization all result in genetically unique individuals
Sexual Reproduction
Sexual reproduction was an early evolutionary innovation after the appearance of eukaryotic cells. It appears to have been very successful because most eukaryotes are able to reproduce sexually, and in many animals, it is the only mode of reproduction. And yet, scientists recognize some real disadvantages to sexual reproduction. On the surface, creating offspring that are genetic clones of the parent appears to be a better system. If the parent organism is successfully occupying a habitat, offspring with the same traits would be similarly successful. There is also the obvious benefit to an organism that can produce offspring whenever circumstances are favorable by asexual budding, fragmentation, or asexual eggs. These methods of reproduction do not require another organism of the opposite sex. Indeed, some organisms that lead a solitary lifestyle have retained the ability to reproduce asexually. In addition, in asexual populations, every individual is capable of reproduction. In sexual populations, the males are not producing the offspring themselves, so in theory an asexual population could grow twice as fast.
However, multicellular organisms that exclusively depend on asexual reproduction are exceedingly rare. Why is sexuality (and meiosis) so common? This is one of the important unanswered questions in biology and has been the focus of much research beginning in the latter half of the twentieth century. There are several possible explanations, one of which is that the variation that sexual reproduction creates among offspring is very important to the survival and reproduction of the population. Thus, on average, a sexually reproducing population will leave more descendants than an otherwise similar asexually reproducing population. The only source of variation in asexual organisms is mutation. This is the ultimate source of variation in sexual organisms, but in addition, those different mutations are continually reshuffled from one generation to the next when different parents combine their unique genomes and the genes are mixed into different combinations by crossovers during prophase I and random assortment at metaphase I.
The Red Queen Hypothesis
It is not in dispute that sexual reproduction provides evolutionary advantages to organisms that employ this mechanism to produce offspring. But why, even in the face of fairly stable conditions, does sexual reproduction persist when it is more difficult and costly for individual organisms? Variation is the outcome of sexual reproduction, but why are ongoing variations necessary? Enter the Red Queen hypothesis, first proposed by Leigh Van Valen in 1973. The concept was named in reference to the Red Queen’s race in Lewis Carroll’s book, Through the Looking-Glass.
All species co-evolve with other organisms; for example predators evolve with their prey, and parasites evolve with their hosts. Each tiny advantage gained by favorable variation gives a species an edge over close competitors, predators, parasites, or even prey. The only method that will allow a co-evolving species to maintain its own share of the resources is to also continually improve its fitness. As one species gains an advantage, this increases selection on the other species; they must also develop an advantage or they will be outcompeted. No single species progresses too far ahead because genetic variation among the progeny of sexual reproduction provides all species with a mechanism to improve rapidly. Species that cannot keep up become extinct. The Red Queen’s catchphrase was, “It takes all the running you can do to stay in the same place.” This is an apt description of co-evolution between competing species.
Life Cycles of Sexually Reproducing Organisms
Fertilization and meiosis alternate in sexual life cycles. What happens between these two events depends on the organism. The process of meiosis reduces the chromosome number by half. Fertilization, the joining of two haploid gametes, restores the diploid condition. There are three main categories of life cycles in multicellular organisms: diploid-dominant, in which the multicellular diploid stage is the most obvious life stage, such as with most animals including humans; haploid-dominant, in which the multicellular haploid stage is the most obvious life stage, such as with all fungi and some algae; and alternation of generations, in which the two stages are apparent to different degrees depending on the group, as with plants and some algae.
Diploid-Dominant Life Cycle
Nearly all animals employ a diploid-dominant life-cycle strategy in which the only haploid cells produced by the organism are the gametes. Early in the development of the embryo, specialized diploid cells, called germ cells, are produced within the gonads, such as the testes and ovaries. Germ cells are capable of mitosis to perpetuate the cell line and meiosis to produce gametes. Once the haploid gametes are formed, they lose the ability to divide again. There is no multicellular haploid life stage. Fertilization occurs with the fusion of two gametes, usually from different individuals, restoring the diploid state (Figure 1).

Figure 1. In animals, sexually reproducing adults form haploid gametes from diploid germ cells. Fusion of the gametes gives rise to a fertilized egg cell, or zygote. The zygote will undergo multiple rounds of mitosis to produce a multicellular offspring. The germ cells are generated early in the development of the zygote.
Haploid-Dominant Life Cycle
Most fungi and algae employ a life-cycle type in which the “body” of the organism—the ecologically important part of the life cycle—is haploid. The haploid cells that make up the tissues of the dominant multicellular stage are formed by mitosis. During sexual reproduction, specialized haploid cells from two individuals, designated the (+) and (−) mating types, join to form a diploid zygote. The zygote immediately undergoes meiosis to form four haploid cells called spores. Although haploid like the “parents,” these spores contain a new genetic combination from two parents. The spores can remain dormant for various time periods. Eventually, when conditions are conducive, the spores form multicellular haploid structures by many rounds of mitosis (Example 1).
Practice Question

Figure 2. Fungi, such as black bread mold (Rhizopus nigricans), have haploid-dominant life cycles. The haploid multicellular stage produces specialized haploid cells by mitosis that fuse to form a diploid zygote. The zygote undergoes meiosis to produce haploid spores. Each spore gives rise to a multicellular haploid organism by mitosis. (credit “zygomycota” micrograph: modification of work by “Fanaberka”/Wikimedia Commons)
If a mutation occurs so that a fungus is no longer able to produce a minus mating type, will it still be able to reproduce?
Alternation of Generations
The third life-cycle type, employed by some algae and all plants, is a blend of the haploid-dominant and diploid-dominant extremes. Species with alternation of generations have both haploid and diploid multicellular organisms as part of their life cycle. The haploid multicellular plants are called gametophytes, because they produce gametes from specialized cells. Meiosis is not directly involved in the production of gametes in this case, because the organism that produces the gametes is already a haploid. Fertilization between the gametes forms a diploid zygote. The zygote will undergo many rounds of mitosis and give rise to a diploid multicellular plant called a sporophyte. Specialized cells of the sporophyte will undergo meiosis and produce haploid spores. The spores will subsequently develop into the gametophytes (Figure 3).

Figure 3. Plants have a life cycle that alternates between a multicellular haploid organism and a multicellular diploid organism. In some plants, such as ferns, both the haploid and diploid plant stages are free-living. The diploid plant is called a sporophyte because it produces haploid spores by meiosis. The spores develop into multicellular, haploid plants called gametophytes because they produce gametes. The gametes of two individuals will fuse to form a diploid zygote that becomes the sporophyte. (credit “fern”: modification of work by Cory Zanker; credit “sporangia”: modification of work by “Obsidian Soul”/Wikimedia Commons; credit “gametophyte and sporophyte”: modification of work by “Vlmastra”/Wikimedia Commons)
Although all plants utilize some version of the alternation of generations, the relative size of the sporophyte and the gametophyte and the relationship between them vary greatly. In plants such as moss, the gametophyte organism is the free-living plant, and the sporophyte is physically dependent on the gametophyte. In other plants, such as ferns, both the gametophyte and sporophyte plants are free-living; however, the sporophyte is much larger. In seed plants, such as magnolia trees and daisies, the gametophyte is composed of only a few cells and, in the case of the female gametophyte, is completely retained within the sporophyte.
Sexual reproduction takes many forms in multicellular organisms. However, at some point in each type of life cycle, meiosis produces haploid cells that will fuse with the haploid cell of another organism. The mechanisms of variation—crossover, random assortment of homologous chromosomes, and random fertilization—are present in all versions of sexual reproduction. The fact that nearly every multicellular organism on Earth employs sexual reproduction is strong evidence for the benefits of producing offspring with unique gene combinations, though there are other possible benefits as well.
Genetic Variation in Meiosis
The gametes produced in meiosis aren’t genetically identical to the starting cell, and they also aren’t identical to one another. As an example, consider the meiosis II diagram above, which shows the end products of meiosis for a simple cell with a diploid number of 2n = 4 chromosomes. The four gametes produced at the end of meiosis II are all slightly different, each with a unique combination of the genetic material present in the starting cell.
As it turns out, there are many more potential gamete types than just the four shown in the diagram, even for a simple cell with with only four chromosomes. This diversity of possible gametes reflects two factors: crossing over and the random orientation of homologue pairs during metaphase of meiosis I.
- Crossing over. The points where homologues cross over and exchange genetic material are chosen more or less at random, and they will be different in each cell that goes through meiosis. If meiosis happens many times, as it does in human ovaries and testes, crossovers will happen at many different points. This repetition produces a wide variety of recombinant chromosomes, chromosomes where fragments of DNA have been exchanged between homologues.
- Random orientation of homologue pairs. The random orientation of homologue pairs during metaphase of meiosis I is another important source of gamete diversity.
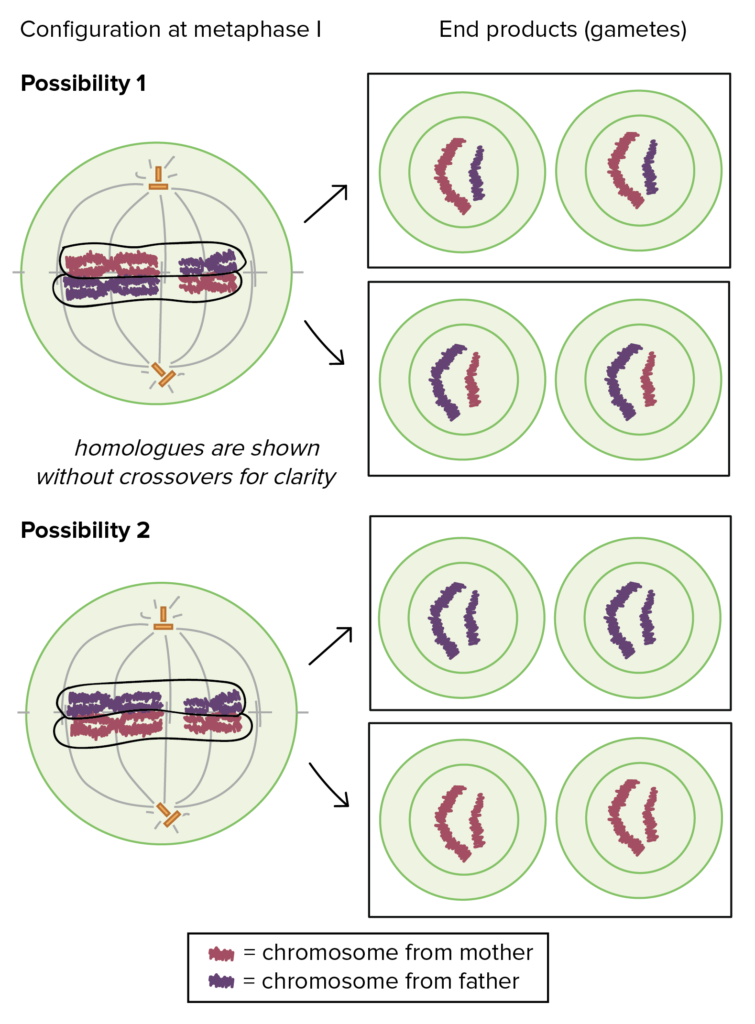
Figure 4. Chromosome configuration and homologue segregation
What exactly does random orientation mean here? Well, a homologous pair consists of one homologue from your dad and one from your mom, and you have 23 pairs of homologous chromosomes all together, counting the X and Y as homologous for this purpose. During meiosis I, the homologous pairs will separate to form two equal groups, but it’s not usually the case that all the paternal—dad—chromosomes will go into one group and all the maternal—mom—chromosomes into the other.
Instead, each pair of homologues will effectively flip a coin to decide which chromosome goes into which group. In a cell with just two pairs of homologous chromosomes, like the one at right, random metaphase orientation allows for 22 = 4 different types of possible gametes. In a human cell, the same mechanism allows for 223 = 8,388,608 different types of possible gametes[1]. And that’s not even considering crossovers!
Given those kinds of numbers, it’s very unlikely that any two sperm or egg cells made by a person will be the same. It’s even more unlikely that you and your sister or brother will be genetically identical, unless you happen to be identical twins, thanks to the process of fertilization (in which a unique egg from Mom combines with a unique sperm from Dad, making a zygote whose genotype is well beyond one-in-a-trillion!)[2].
Meiosis and fertilization create genetic variation by making new combinations of gene variants (alleles). In some cases, these new combinations may make an organism more or less fit (able to survive and reproduce), thus providing the raw material for natural selection. Genetic variation is important in allowing a population to adapt via natural selection and thus survive in the long term.
Chromosomal Structural Rearrangements
Cytologists have characterized numerous structural rearrangements in chromosomes, but chromosome inversions and translocations are the most common. Both are identified during meiosis by the adaptive pairing of rearranged chromosomes with their former homologs to maintain appropriate gene alignment. If the genes carried on two homologs are not oriented correctly, a recombination event could result in the loss of genes from one chromosome and the gain of genes on the other. This would produce aneuploid gametes.
Chromosome Inversions
A chromosome inversion is the detachment, 180° rotation, and reinsertion of part of a chromosome. Inversions may occur in nature as a result of mechanical shear, or from the action of transposable elements (special DNA sequences capable of facilitating the rearrangement of chromosome segments with the help of enzymes that cut and paste DNA sequences). Unless they disrupt a gene sequence, inversions only change the orientation of genes and are likely to have more mild effects than aneuploid errors. However, altered gene orientation can result in functional changes because regulators of gene expression could be moved out of position with respect to their targets, causing aberrant levels of gene products.
An inversion can be pericentric and include the centromere, or paracentric and occur outside of the centromere (Figure 5). A pericentric inversion that is asymmetric about the centromere can change the relative lengths of the chromosome arms, making these inversions easily identifiable.

Figure 5. Pericentric inversions include the centromere, and paracentric inversions do not. A pericentric inversion can change the relative lengths of the chromosome arms; a paracentric inversion cannot.
When one homologous chromosome undergoes an inversion but the other does not, the individual is described as an inversion heterozygote. To maintain point-for-point synapsis during meiosis, one homolog must form a loop, and the other homolog must mold around it. Although this topology can ensure that the genes are correctly aligned, it also forces the homologs to stretch and can be associated with regions of imprecise synapsis (Figure 6).

Figure 6. When one chromosome undergoes an inversion but the other does not, one chromosome must form an inverted loop to retain point-for-point interaction during synapsis. This inversion pairing is essential to maintaining gene alignment during meiosis and to allow for recombination.
The Chromosome 18 Inversion
Not all structural rearrangements of chromosomes produce nonviable, impaired, or infertile individuals. In rare instances, such a change can result in the evolution of a new species. In fact, a pericentric inversion in chromosome 18 appears to have contributed to the evolution of humans. This inversion is not present in our closest genetic relatives, the chimpanzees. Humans and chimpanzees differ cytogenetically by pericentric inversions on several chromosomes and by the fusion of two separate chromosomes in chimpanzees that correspond to chromosome two in humans.
The pericentric chromosome 18 inversion is believed to have occurred in early humans following their divergence from a common ancestor with chimpanzees approximately five million years ago. Researchers characterizing this inversion have suggested that approximately 19,000 nucleotide bases were duplicated on 18p, and the duplicated region inverted and reinserted on chromosome 18 of an ancestral human.
A comparison of human and chimpanzee genes in the region of this inversion indicates that two genes—ROCK1 and USP14—that are adjacent on chimpanzee chromosome 17 (which corresponds to human chromosome 18) are more distantly positioned on human chromosome 18. This suggests that one of the inversion breakpoints occurred between these two genes. Interestingly, humans and chimpanzees express USP14 at distinct levels in specific cell types, including cortical cells and fibroblasts. Perhaps the chromosome 18 inversion in an ancestral human repositioned specific genes and reset their expression levels in a useful way. Because both ROCK1 and USP14 encode cellular enzymes, a change in their expression could alter cellular function. It is not known how this inversion contributed to hominid evolution, but it appears to be a significant factor in the divergence of humans from other primates.
Translocations
A translocation occurs when a segment of a chromosome dissociates and reattaches to a different, nonhomologous chromosome. Translocations can be benign or have devastating effects depending on how the positions of genes are altered with respect to regulatory sequences. Notably, specific translocations have been associated with several cancers and with schizophrenia. Reciprocal translocations result from the exchange of chromosome segments between two nonhomologous chromosomes such that there is no gain or loss of genetic information (Figure 7).

Figure 7. A reciprocal translocation occurs when a segment of DNA is transferred from one chromosome to another, nonhomologous chromosome. (credit: modification of work by National Human Genome Research/USA)
Video Review: Genetic Diversity
As you now know, genetic variation is very important. Genetic variation is introduced in multiple ways, including changes in mitosis, crossing over and random orientation in meiosis, and random fertilization. The video below offers you a nice overview of how each contributes to genetic diversity.
Check Your Understanding
Answer the question(s) below to see how well you understand the topics covered in the previous section. This short quiz does not count toward your grade in the class, and you can retake it an unlimited number of times.
Use this quiz to check your understanding and decide whether to (1) study the previous section further or (2) move on to the next section.
Candela Citations
- Introduction to Genetic Diversity. Authored by: Shelli Carter and Lumen Learning. Provided by: Lumen Learning. License: CC BY: Attribution
- Biology. Provided by: OpenStax CNX. Located at: http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8. License: CC BY: Attribution. License Terms: Download for free at http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8
- Genetic Variation in Meiosis. Provided by: Khan Academy. Project: https://www.khanacademy.org/science/biology/cellular-molecular-biology/meiosis/a/phases-of-meiosis. License: CC BY-NC-SA: Attribution-NonCommercial-ShareAlike
- Mitosis, Meiosis and Sexual Reproduction. Authored by: Khan Academy. Located at: https://youtu.be/kaSIjIzAtYA. License: All Rights Reserved. License Terms: Standard YouTube License