Discuss the scope and study of community ecology
Populations rarely, if ever, live in isolation from populations of other species. In most cases, numerous species share a habitat. The interactions between these populations play a major role in regulating population growth and abundance. All populations occupying the same habitat form a community: populations inhabiting a specific area at the same time. The number of species occupying the same habitat and their relative abundance is known as species diversity. Areas with low diversity, such as the glaciers of Antarctica, still contain a wide variety of living things, whereas the diversity of tropical rainforests is so great that it cannot be counted. Ecology is studied at the community level to understand how species interact with each other and compete for the same resources.
Learning Objectives
- Discuss the importance of predation and herbivory in the ecosystem, including how organisms defend against these
- Define the competitive exclusion principle
- Compare and contrast the three different types of symbiotic relationships
- Identify different species roles that structure communities
- Describe community dynamics as the changes in community structure that take place over time
- Define behavioral biology
- Identify different types of innate behaviors in animals
- Identify different types of learned behaviors in animals
Predation and Herbivory
Perhaps the classical example of species interaction is predation: the hunting of prey by its predator. Nature shows on television highlight the drama of one living organism killing another. Populations of predators and prey in a community are not constant over time: in most cases, they vary in cycles that appear to be related. The most often cited example of predator-prey dynamics is seen in the cycling of the lynx (predator) and the snowshoe hare (prey), using nearly 200 year-old trapping data from North American forests (Figure 1). This cycle of predator and prey lasts approximately 10 years, with the predator population lagging 1–2 years behind that of the prey population. As the hare numbers increase, there is more food available for the lynx, allowing the lynx population to increase as well. When the lynx population grows to a threshold level, however, they kill so many hares that hare population begins to decline, followed by a decline in the lynx population because of scarcity of food. When the lynx population is low, the hare population size begins to increase due, at least in part, to low predation pressure, starting the cycle anew.

Figure 1. The cycling of lynx and snowshoe hare populations in Northern Ontario is an example of predator-prey dynamics.
The idea that the population cycling of the two species is entirely controlled by predation models has come under question. More recent studies have pointed to undefined density-dependent factors as being important in the cycling, in addition to predation. One possibility is that the cycling is inherent in the hare population due to density-dependent effects such as lower fecundity (maternal stress) caused by crowding when the hare population gets too dense. The hare cycling would then induce the cycling of the lynx because it is the lynxes’ major food source. The more we study communities, the more complexities we find, allowing ecologists to derive more accurate and sophisticated models of population dynamics.
Herbivory describes the consumption of plants by insects and other animals, and it is another interspecific relationship that affects populations. Unlike animals, most plants cannot outrun predators or use mimicry to hide from hungry animals. Some plants have developed mechanisms to defend against herbivory. Other species have developed mutualistic relationships; for example, herbivory provides a mechanism of seed distribution that aids in plant reproduction.
Defense Mechanisms against Predation and Herbivory
The study of communities must consider evolutionary forces that act on the members of the various populations contained within it. Species are not static, but slowly changing and adapting to their environment by natural selection and other evolutionary forces. Species have evolved numerous mechanisms to escape predation and herbivory. These defenses may be mechanical, chemical, physical, or behavioral.
Mechanical defenses, such as the presence of thorns on plants or the hard shell on turtles, discourage animal predation and herbivory by causing physical pain to the predator or by physically preventing the predator from being able to eat the prey. Chemical defenses are produced by many animals as well as plants, such as the foxglove which is extremely toxic when eaten. Figure 2 shows some organisms’ defenses against predation and herbivory.

Figure 2. The (a) honey locust tree (Gleditsia triacanthos) uses thorns, a mechanical defense, against herbivores, while the (b) Florida red-bellied turtle (Pseudemys nelsoni) uses its shell as a mechanical defense against predators. (c) Foxglove (Digitalis sp.) uses a chemical defense: toxins produced by the plant can cause nausea, vomiting, hallucinations, convulsions, or death when consumed. (d) The North American millipede (Narceus americanus) uses both mechanical and chemical defenses: when threatened, the millipede curls into a defensive ball and produces a noxious substance that irritates eyes and skin. (credit a: modification of work by Huw Williams; credit b: modification of work by “JamieS93”/Flickr; credit c: modification of work by Philip Jägenstedt; credit d: modification of work by Cory Zanker)
Many species use their body shape and coloration to avoid being detected by predators. The tropical walking stick is an insect with the coloration and body shape of a twig which makes it very hard to see when stationary against a background of real twigs (Figure 3a). In another example, the chameleon can change its color to match its surroundings (Figure 3b). Both of these are examples of camouflage, or avoiding detection by blending in with the background.

Figure 3. (a) The tropical walking stick and (b) the chameleon use body shape and/or coloration to prevent detection by predators. (credit a: modification of work by Linda Tanner; credit b: modification of work by Frank Vassen)
Some species use coloration as a way of warning predators that they are not good to eat. For example, the cinnabar moth caterpillar, the fire-bellied toad, and many species of beetle have bright colors that warn of a foul taste, the presence of toxic chemical, and/or the ability to sting or bite, respectively. Predators that ignore this coloration and eat the organisms will experience their unpleasant taste or presence of toxic chemicals and learn not to eat them in the future. This type of defensive mechanism is called aposematic coloration, or warning coloration.

Figure 4. (a) The strawberry poison dart frog (Oophaga pumilio) uses aposematic coloration to warn predators that it is toxic, while the (b) striped skunk (Mephitis mephitis) uses aposematic coloration to warn predators of the unpleasant odor it produces. (credit a: modification of work by Jay Iwasaki; credit b: modification of work by Dan Dzurisin)
While some predators learn to avoid eating certain potential prey because of their coloration, other species have evolved mechanisms to mimic this coloration to avoid being eaten, even though they themselves may not be unpleasant to eat or contain toxic chemicals. In Batesian mimicry, a harmless species imitates the warning coloration of a harmful one. Assuming they share the same predators, this coloration then protects the harmless ones, even though they do not have the same level of physical or chemical defenses against predation as the organism they mimic. Many insect species mimic the coloration of wasps or bees, which are stinging, venomous insects, thereby discouraging predation (Figure 5).

Figure 5. Batesian mimicry occurs when a harmless species mimics the coloration of a harmful species, as is seen with the (a) bumblebee and (b) bee-like robber fly. (credit a, b: modification of work by Cory Zanker)

Figure 6. Several unpleasant-tasting Heliconius butterfly species share a similar color pattern with better-tasting varieties. (credit: Joron M, Papa R, Beltrán M, Chamberlain N, Mavárez J, et al.)
In Müllerian mimicry, multiple species share the same warning coloration, but all of them actually have defenses. Figure 6 shows a variety of foul-tasting butterflies with similar coloration.
In Emsleyan/Mertensian mimicry, a deadly prey mimics a less dangerous one, such as the venomous coral snake mimicking the non-venomous milk snake. This type of mimicry is extremely rare and more difficult to understand than the previous two types. For this type of mimicry to work, it is essential that eating the milk snake has unpleasant, but not fatal, consequences. Then, these predators learn not to eat snakes with this coloration, protecting the coral snake as well. If the snake were fatal to the predator, there would be no opportunity for the predator to learn not to eat it, and the benefit for the less toxic species would disappear.
Competition
Resources are often limited within a habitat and multiple species may compete to obtain them. All species have an ecological niche in the ecosystem, which describes how they acquire the resources they need and how they interact with other species in the community. The competitive exclusion principle states that two species cannot occupy the same niche in a habitat. In other words, different species cannot coexist in a community if they are competing for all the same resources. An example of this principle is shown in Figure 7, with two protozoan species, Paramecium aurelia and Paramecium caudatum. When grown individually in the laboratory, they both thrive. But when they are placed together in the same test tube (habitat), P. aurelia outcompetes P. caudatum for food, leading to the latter’s eventual extinction.

Figure 7. Paramecium aurelia and Paramecium caudatum grow well individually, but when they compete for the same resources, the P. aurelia outcompetes the P. caudatum.
Resource Partitioning
Competitive exclusion may be avoided if one or both of the competing species evolves to use a different resource, occupy a different area of the habitat, or feed during a different time of day. The result of this kind of evolution is that two similar species use largely non-overlapping resources and thus have different niches. This is called resource partitioning, and it helps the species coexist because there is less direct competition between them.
The anole lizards found on the island of Puerto Rico are a good example of resource partitioning. In this group, natural selection has led to the evolution of different species that make use of different resources. Figure 8 shows resource partitioning among 11 species of anole lizards. Each species lives in its own preferred habitat, which is defined by type and height of vegetation (trees, shrubs, cactus, etc.), sunlight, and moisture, among other factors.

Figure 8. Resource partitioning among anole lizards
Video Review
Watch this video to review competition and how populations share resources in a community:
Symbiosis
Symbiotic relationships, or symbioses (plural), are close interactions between individuals of different species over an extended period of time which impact the abundance and distribution of the associating populations. Most scientists accept this definition, but some restrict the term to only those species that are mutualistic, where both individuals benefit from the interaction. In this discussion, the broader definition will be used.
Commensalism

Figure 9. The southern masked-weaver bird is starting to make a nest in a tree in Zambezi Valley, Zambia. This is an example of a commensal relationship, in which one species (the bird) benefits, while the other (the tree) neither benefits nor is harmed. (credit: “Hanay”/Wikimedia Commons)
A commensal relationship occurs when one species benefits from the close, prolonged interaction, while the other neither benefits nor is harmed. Birds nesting in trees provide an example of a commensal relationship (Figure 9). The tree is not harmed by the presence of the nest among its branches. The nests are light and produce little strain on the structural integrity of the branch, and most of the leaves, which the tree uses to get energy by photosynthesis, are above the nest so they are unaffected. The bird, on the other hand, benefits greatly. If the bird had to nest in the open, its eggs and young would be vulnerable to predators. Another example of a commensal relationship is the clown fish and the sea anemone. The sea anemone is not harmed by the fish, and the fish benefits with protection from predators who would be stung upon nearing the sea anemone.
Mutualism
A second type of symbiotic relationship is called mutualism, where two species benefit from their interaction. Some scientists believe that these are the only true examples of symbiosis. For example, termites have a mutualistic relationship with protozoa that live in the insect’s gut (Figure 10a). The termite benefits from the ability of bacterial symbionts within the protozoa to digest cellulose. The termite itself cannot do this, and without the protozoa, it would not be able to obtain energy from its food (cellulose from the wood it chews and eats). The protozoa and the bacterial symbionts benefit by having a protective environment and a constant supply of food from the wood chewing actions of the termite. Lichens have a mutualistic relationship between fungus and photosynthetic algae or bacteria (Figure 10b). As these symbionts grow together, the glucose produced by the algae provides nourishment for both organisms, whereas the physical structure of the lichen protects the algae from the elements and makes certain nutrients in the atmosphere more available to the algae.

Figure 10. (a) Termites form a mutualistic relationship with symbiotic protozoa in their guts, which allow both organisms to obtain energy from the cellulose the termite consumes. (b) Lichen is a fungus that has symbiotic photosynthetic algae living inside its cells. (credit a: modification of work by Scott Bauer, USDA; credit b: modification of work by Cory Zanker)
Parasitism
A parasite is an organism that lives in or on another living organism and derives nutrients from it. In this relationship, the parasite benefits, but the organism being fed upon, the host is harmed. The host is usually weakened by the parasite as it siphons resources the host would normally use to maintain itself. The parasite, however, is unlikely to kill the host, especially not quickly, because this would allow no time for the organism to complete its reproductive cycle by spreading to another host.
The reproductive cycles of parasites are often very complex, sometimes requiring more than one host species. A tapeworm is a parasite that causes disease in humans when contaminated, undercooked meat such as pork, fish, or beef is consumed (Figure 11). The tapeworm can live inside the intestine of the host for several years, benefiting from the food the host is bringing into its gut by eating, and may grow to be over 50 ft long by adding segments. The parasite moves from species to species in a cycle, making two hosts necessary to complete its life cycle. Another common parasite is Plasmodium falciparum, the protozoan cause of malaria, a significant disease in many parts of the world. Living in human liver and red blood cells, the organism reproduces asexually in the gut of blood-feeding mosquitoes to complete its life cycle. Thus malaria is spread from human to human by mosquitoes, one of many arthropod-borne infectious diseases.

Figure 11. This diagram shows the life cycle of a pork tapeworm (Taenia solium), a human worm parasite. (credit: modification of work by CDC)
Community Structure
Communities are complex entities that can be characterized by their structure (the types and numbers of species present) and dynamics (how communities change over time). Understanding community structure and dynamics enables community ecologists to manage ecosystems more effectively.
Foundation Species

Figure 12. Coral is the foundation species of coral reef ecosystems. (credit: Jim E. Maragos, USFWS)
Foundation species are considered the “base” or “bedrock” of a community, having the greatest influence on its overall structure. They are usually the primary producers: organisms that bring most of the energy into the community. Kelp, brown algae, is a foundation species, forming the basis of the kelp forests off the coast of California.
Foundation species may physically modify the environment to produce and maintain habitats that benefit the other organisms that use them. An example is the photosynthetic corals of the coral reef (Figure 12). Corals themselves are not photosynthetic, but harbor symbionts within their body tissues (dinoflagellates called zooxanthellae) that perform photosynthesis; this is another example of a mutualism. The exoskeletons of living and dead coral make up most of the reef structure, which protects many other species from waves and ocean currents.
Biodiversity, Species Richness, and Relative Species Abundance
Biodiversity describes a community’s biological complexity: it is measured by the number of different species (species richness) in a particular area and their relative abundance (species evenness). The area in question could be a habitat, a biome, or the entire biosphere. Species richness is the term that is used to describe the number of species living in a habitat or biome. Species richness varies across the globe (Figure 13). One factor in determining species richness is latitude, with the greatest species richness occurring in ecosystems near the equator, which often have warmer temperatures, large amounts of rainfall, and low seasonality. The lowest species richness occurs near the poles, which are much colder, drier, and thus less conducive to life in Geologic time (time since glaciations). The predictability of climate or productivity is also an important factor. Other factors influence species richness as well. For example, the study of island biogeography attempts to explain the relatively high species richness found in certain isolated island chains, including the Galápagos Islands that inspired the young Darwin. Relative species abundance is the number of individuals in a species relative to the total number of individuals in all species within a habitat, ecosystem, or biome. Foundation species often have the highest relative abundance of species.

Figure 13. The greatest species richness for mammals in North and South America is associated with the equatorial latitudes. (credit: modification of work by NASA, CIESIN, Columbia University)
Keystone Species

Figure 14. The Pisaster ochraceus sea star is a keystone species. (credit: Jerry Kirkhart)
A keystone species is one whose presence is key to maintaining biodiversity within an ecosystem and to upholding an ecological community’s structure. The intertidal sea star, Pisaster ochraceus, of the northwestern United States is a keystone species (Figure 14).
Studies have shown that when this organism is removed from communities, populations of their natural prey (mussels) increase, completely altering the species composition and reducing biodiversity. Another keystone species is the banded tetra, a fish in tropical streams, which supplies nearly all of the phosphorus, a necessary inorganic nutrient, to the rest of the community. If these fish were to become extinct, the community would be greatly affected.
Invasive Species

Figure 15. Aquatic invasive species in the United States: (a) purple loosestrife and (b) zebra mussel. (credit a: modification of work by Liz West; credit b: modification of work by M. McCormick, NOAA)
Invasive species are non-native organisms that, when introduced to an area out of their native range, threaten the ecosystem balance of that habitat. Many such species exist in the United States, as shown in Figures 4–6. Whether enjoying a forest hike, taking a summer boat trip, or simply walking down an urban street, you have likely encountered an invasive species.
Invasive species like purple loosestrife (Lythrum salicaria) and the zebra mussel (Dreissena polymorpha) threaten certain aquatic ecosystems (Figure 15).
Some forests are threatened by the spread of common buckthorn (Rhamnus cathartica), garlic mustard (Alliaria petiolata), and the emerald ash borer (Agrilus planipennis) (Figure 16). The European starling (Sturnus vulgaris) may compete with native bird species for nest holes.

Figure 16. Invasive species in US forests: (a) common buckthorn, (b) garlic mustard, and (c) the emerald ash borer. (credit a: modification of work by E. Dronkert; credit b: modification of work by Dan Davison; credit c: modification of work by USDA)
Community Dynamics
Community dynamics are the changes in community structure and composition over time. Sometimes these changes are induced by environmental disturbances such as volcanoes, earthquakes, storms, fires, and climate change. Communities with a stable structure are said to be at equilibrium. Following a disturbance, the community may or may not return to the equilibrium state.
Succession describes the sequential appearance and disappearance of species in a community over time. In primary succession, newly exposed or newly formed land is colonized by living things; in secondary succession, part of an ecosystem is disturbed and remnants of the previous community remain.
Primary Succession and Pioneer Species

Figure 17. During primary succession in lava on Maui, Hawaii, succulent plants are the pioneer species. (credit: Forest and Kim Starr)
Primary succession occurs when new land is formed or rock is exposed: for example, following the eruption of volcanoes, such as those on the Big Island of Hawaii. As lava flows into the ocean, new land is continually being formed. On the Big Island, approximately 32 acres of land is added each year. First, weathering and other natural forces break down the substrate enough for the establishment of certain hearty plants and lichens with few soil requirements, known as pioneer species (Figure 17). These species help to further break down the mineral rich lava into soil where other, less hardy species will grow and eventually replace the pioneer species. In addition, as these early species grow and die, they add to an ever-growing layer of decomposing organic material and contribute to soil formation. Over time the area will reach an equilibrium state, with a set of organisms quite different from the pioneer species.
Secondary Succession
A classic example of secondary succession occurs in oak and hickory forests cleared by wildfire (Figure 18). Wildfires will burn most vegetation and kill those animals unable to flee the area. Their nutrients, however, are returned to the ground in the form of ash. Thus, even when areas are devoid of life due to severe fires, the area will soon be ready for new life to take hold.
Before the fire, the vegetation was dominated by tall trees with access to the major plant energy resource: sunlight. Their height gave them access to sunlight while also shading the ground and other low-lying species. After the fire, though, these trees are no longer dominant. Thus, the first plants to grow back are usually annual plants followed within a few years by quickly growing and spreading grasses and other pioneer species. Due to, at least in part, changes in the environment brought on by the growth of the grasses and other species, over many years, shrubs will emerge along with small pine, oak, and hickory trees. These organisms are called intermediate species. Eventually, over 150 years, the forest will reach its equilibrium point where species composition is no longer changing and resembles the community before the fire. This equilibrium state is referred to as the climax community, which will remain stable until the next disturbance.

Figure 18. Secondary succession is shown in an oak and hickory forest after a forest fire.
Behavioral Biology
Behavior is the change in activity of an organism in response to a stimulus. Behavioral biology is the study of the biological and evolutionary bases for such changes. The idea that behaviors evolved as a result of the pressures of natural selection is not new. Animal behavior has been studied for decades, by biologists in the science of ethology, by psychologists in the science of comparative psychology, and by scientists of many disciplines in the study of neurobiology. Although there is overlap between these disciplines, scientists in these behavioral fields take different approaches. Comparative psychology is an extension of work done in human and behavioral psychology. Ethology is an extension of genetics, evolution, anatomy, physiology, and other biological disciplines. Still, one cannot study behavioral biology without touching on both comparative psychology and ethology.
Innate Behaviors
One goal of behavioral biology is to dissect out the innate behaviors, which have a strong genetic component and are largely independent of environmental influences, from the learned behaviors, which result from environmental conditioning. Innate behavior, or instinct, is important because there is no risk of an incorrect behavior being learned. They are “hard wired” into the system. On the other hand, learned behaviors, although riskier, are flexible, dynamic, and can be altered according to changes in the environment.
Movement and Migration
Innate or instinctual behaviors rely on response to stimuli. The simplest example of this is a reflex action, an involuntary and rapid response to stimulus. To test the “knee-jerk” reflex, a doctor taps the patellar tendon below the kneecap with a rubber hammer. The stimulation of the nerves there leads to the reflex of extending the leg at the knee. This is similar to the reaction of someone who touches a hot stove and instinctually pulls his or her hand away. Even humans, with our great capacity to learn, still exhibit a variety of innate behaviors.
Kinesis and Taxis
Another activity or movement of innate behavior is kinesis, or the undirected movement in response to a stimulus. Orthokinesis is the increased or decreased speed of movement of an organism in response to a stimulus. Woodlice, for example, increase their speed of movement when exposed to high or low temperatures. This movement, although random, increases the probability that the insect spends less time in the unfavorable environment. Another example is klinokinesis, an increase in turning behaviors. It is exhibited by bacteria such as E. coli which, in association with orthokinesis, helps the organisms randomly find a more hospitable environment.
A similar, but more directed version of kinesis is taxis: the directed movement towards or away from a stimulus. This movement can be in response to light (phototaxis), chemical signals (chemotaxis), or gravity (geotaxis) and can be directed toward (positive) or away (negative) from the source of the stimulus. An example of a positive chemotaxis is exhibited by the unicellular protozoan Tetrahymena thermophila. This organism swims using its cilia, at times moving in a straight line, and at other times making turns. The attracting chemotactic agent alters the frequency of turning as the organism moves directly toward the source, following the increasing concentration gradient.
Fixed Action Patterns
A fixed action pattern is a series of movements elicited by a stimulus such that even when the stimulus is removed, the pattern goes on to completion. An example of such a behavior occurs in the three-spined stickleback, a small freshwater fish (Figure 19). Males of this species develop a red belly during breeding season and show instinctual aggressiveness to other males during this time. In laboratory experiments, researchers exposed such fish to objects that in no way resemble a fish in their shape, but which were painted red on their lower halves. The male sticklebacks responded aggressively to the objects just as if they were real male sticklebacks.

Figure 19. Male three-spined stickleback fish exhibit a fixed action pattern. During mating season, the males, which develop a bright red belly, react strongly to red-bottomed objects that in no way resemble fish.
Migration

Figure 20. Wildebeests migrate in a clockwise fashion over 1800 miles each year in search of rain-ripened grass. (credit: Eric Inafuku)
Migration is the long-range seasonal movement of animals. It is an evolved, adapted response to variation in resource availability, and it is a common phenomenon found in all major groups of animals. Birds fly south for the winter to get to warmer climates with sufficient food, and salmon migrate to their spawning grounds. The popular 2005 documentary March of the Penguins followed the 62-mile migration of emperor penguins through Antarctica to bring food back to their breeding site and to their young. Wildebeests (Figure 20) migrate over 1800 miles each year in search of new grasslands.
Although migration is thought of as innate behavior, only some migrating species always migrate (obligate migration). Animals that exhibit facultative migration can choose to migrate or not. Additionally, in some animals, only a portion of the population migrates, whereas the rest does not migrate (incomplete migration). For example, owls that live in the tundra may migrate in years when their food source, small rodents, is relatively scarce, but not migrate during the years when rodents are plentiful.
Foraging

Figure 21. The painted stork uses its long beak to forage. (credit: J.M. Garg)
Foraging is the act of searching for and exploiting food resources. Feeding behaviors that maximize energy gain and minimize energy expenditure are called optimal foraging behaviors, and these are favored by natural section. The painted stork, for example, uses its long beak to search the bottom of a freshwater marshland for crabs and other food (Figure 21).
Living in Groups
Not all animals live in groups, but even those that live relatively solitary lives, with the exception of those that can reproduce asexually, must mate. Mating usually involves one animal signaling another so as to communicate the desire to mate. There are several types of energy-intensive behaviors or displays associated with mating, called mating rituals. Other behaviors found in populations that live in groups are described in terms of which animal benefits from the behavior. In selfish behavior, only the animal in question benefits; in altruistic behavior, one animal’s actions benefit another animal; cooperative behavior describes when both animals benefit. All of these behaviors involve some sort of communication between population members.
Communication within a Species
Animals communicate with each other using stimuli known as signals. An example of this is seen in the three-spined stickleback, where the visual signal of a red region in the lower half of a fish signals males to become aggressive and signals females to mate. Other signals are chemical (pheromones), aural (sound), visual (courtship and aggressive displays), or tactile (touch). These types of communication may be instinctual or learned or a combination of both. These are not the same as the communication we associate with language, which has been observed only in humans and perhaps in some species of primates and cetaceans.
A pheromone is a secreted chemical signal used to obtain a response from another individual of the same species. The purpose of pheromones is to elicit a specific behavior from the receiving individual. Pheromones are especially common among social insects, but they are used by many species to attract the opposite sex, to sound alarms, to mark food trails, and to elicit other, more complex behaviors. Even humans are thought to respond to certain pheromones called axillary steroids. These chemicals influence human perception of other people, and in one study were responsible for a group of women synchronizing their menstrual cycles. The role of pheromones in human-to-human communication is still somewhat controversial and continues to be researched.
Songs are an example of an aural signal, one that needs to be heard by the recipient. Perhaps the best known of these are songs of birds, which identify the species and are used to attract mates. Other well-known songs are those of whales, which are of such low frequency that they can travel long distances underwater. Dolphins communicate with each other using a wide variety of vocalizations. Male crickets make chirping sounds using a specialized organ to attract a mate, repel other males, and to announce a successful mating.

Figure 22. This stork’s courtship display is designed to attract potential mates. (credit: Linda “jinterwas”/Flickr)
Courtship displays are a series of ritualized visual behaviors (signals) designed to attract and convince a member of the opposite sex to mate. These displays are ubiquitous in the animal kingdom. Often these displays involve a series of steps, including an initial display by one member followed by a response from the other. If at any point, the display is performed incorrectly or a proper response is not given, the mating ritual is abandoned and the mating attempt will be unsuccessful. The mating display of the common stork is shown in Figure 22.
Aggressive displays are also common in the animal kingdom. An example is when a dog bares its teeth when it wants another dog to back down. Presumably, these displays communicate not only the willingness of the animal to fight, but also its fighting ability. Although these displays do signal aggression on the part of the sender, it is thought that these displays are actually a mechanism to reduce the amount of actual fighting that occurs between members of the same species: they allow individuals to assess the fighting ability of their opponent and thus decide whether it is “worth the fight.” The testing of certain hypotheses using game theory has led to the conclusion that some of these displays may overstate an animal’s actual fighting ability and are used to “bluff” the opponent. This type of interaction, even if “dishonest,” would be favored by natural selection if it is successful more times than not.
Distraction displays are seen in birds and some fish. They are designed to attract a predator away from the nest that contains their young. This is an example of an altruistic behavior: it benefits the young more than the individual performing the display, which is putting itself at risk by doing so.
Many animals, especially primates, communicate with other members in the group through touch. Activities such as grooming, touching the shoulder or root of the tail, embracing, lip contact, and greeting ceremonies have all been observed in the Indian langur, an Old World monkey. Similar behaviors are found in other primates, especially in the great apes.
The killdeer bird distracts predators from its eggs by faking a broken wing display in this video taken in Boise, Idaho. Note that this video has no narration.
Altruistic Behaviors
Behaviors that lower the fitness of the individual but increase the fitness of another individual are termed altruistic. Examples of such behaviors are seen widely across the animal kingdom. Social insects such as worker bees have no ability to reproduce, yet they maintain the queen so she can populate the hive with her offspring. Meerkats keep a sentry standing guard to warn the rest of the colony about intruders, even though the sentry is putting itself at risk. Wolves and wild dogs bring meat to pack members not present during a hunt. Lemurs take care of infants unrelated to them. Although on the surface, these behaviors appear to be altruistic, it may not be so simple.
There has been much discussion over why altruistic behaviors exist. Do these behaviors lead to overall evolutionary advantages for their species? Do they help the altruistic individual pass on its own genes? And what about such activities between unrelated individuals? One explanation for altruistic-type behaviors is found in the genetics of natural selection. In the 1976 book, The Selfish Gene, scientist Richard Dawkins attempted to explain many seemingly altruistic behaviors from the viewpoint of the gene itself. Although a gene obviously cannot be selfish in the human sense, it may appear that way if the sacrifice of an individual benefits related individuals that share genes that are identical by descent (present in relatives because of common lineage). Mammal parents make this sacrifice to take care of their offspring. Emperor penguins migrate miles in harsh conditions to bring food back for their young. Selfish gene theory has been controversial over the years and is still discussed among scientists in related fields.
Even less-related individuals, those with less genetic identity than that shared by parent and offspring, benefit from seemingly altruistic behavior. The activities of social insects such as bees, wasps, ants, and termites are good examples. Sterile workers in these societies take care of the queen because they are closely related to it, and as the queen has offspring, she is passing on genes from the workers indirectly. Thus, it is of fitness benefit for the worker to maintain the queen without having any direct chance of passing on its genes due to its sterility. The lowering of individual fitness to enhance the reproductive fitness of a relative and thus one’s inclusive fitness evolves through kin selection. This phenomenon can explain many superficially altruistic behaviors seen in animals. However, these behaviors may not be truly defined as altruism in these cases because the actor is actually increasing its own fitness either directly (through its own offspring) or indirectly (through the inclusive fitness it gains through relatives that share genes with it).
Unrelated individuals may also act altruistically to each other, and this seems to defy the “selfish gene” explanation. An example of this observed in many monkey species where a monkey will present its back to an unrelated monkey to have that individual pick the parasites from its fur. After a certain amount of time, the roles are reversed and the first monkey now grooms the second monkey. Thus, there is reciprocity in the behavior. Both benefit from the interaction and their fitness is raised more than if neither cooperated nor if one cooperated and the other did not cooperate. This behavior is still not necessarily altruism, as the “giving” behavior of the actor is based on the expectation that it will be the “receiver” of the behavior in the future, termed reciprocal altruism. Reciprocal altruism requires that individuals repeatedly encounter each other, often the result of living in the same social group, and that cheaters (those that never “give back”) are punished.
Evolutionary game theory, a modification of classical game theory in mathematics, has shown that many of these so-called “altruistic behaviors” are not altruistic at all. The definition of “pure” altruism, based on human behavior, is an action that benefits another without any direct benefit to oneself. Most of the behaviors previously described do not seem to satisfy this definition, and game theorists are good at finding “selfish” components in them. Others have argued that the terms “selfish” and “altruistic” should be dropped completely when discussing animal behavior, as they describe human behavior and may not be directly applicable to instinctual animal activity. What is clear, though, is that heritable behaviors that improve the chances of passing on one’s genes or a portion of one’s genes are favored by natural selection and will be retained in future generations as long as those behaviors convey a fitness advantage. These instinctual behaviors may then be applied, in special circumstances, to other species, as long as it doesn’t lower the animal’s fitness.
Finding Sex Partners
Not all animals reproduce sexually, but many that do have the same challenge: they need to find a suitable mate and often have to compete with other individuals to obtain one. Significant energy is spent in the process of locating, attracting, and mating with the sex partner. Two types of selection occur during this process and can lead to traits that are important to reproduction called secondary sexual characteristics: intersexual selection, the choosing of a mate where individuals of one sex choose mates of the other sex, and intrasexual selection, the competition for mates between species members of the same sex. Intersexual selection is often complex because choosing a mate may be based on a variety of visual, aural, tactile, and chemical cues. An example of intersexual selection is when female peacocks choose to mate with the male with the brightest plumage. This type of selection often leads to traits in the chosen sex that do not enhance survival, but are those traits most attractive to the opposite sex (often at the expense of survival). Intrasexual selection involves mating displays and aggressive mating rituals such as rams butting heads—the winner of these battles is the one that is able to mate. Many of these rituals use up considerable energy but result in the selection of the healthiest, strongest, and/or most dominant individuals for mating. Three general mating systems, all involving innate as opposed to learned behaviors, are seen in animal populations: monogamous, polygynous, and polyandrous.
Watch this informative video on sexual selection.
In monogamous systems, one male and one female are paired for at least one breeding season. In some animals, such as the gray wolf, these associations can last much longer, even a lifetime. Several explanations have been proposed for this type of mating system. The “mate-guarding hypothesis” states that males stay with the female to prevent other males from mating with her. This behavior is advantageous in such situations where mates are scarce and difficult to find. Another explanation is the “male-assistance hypothesis,” where males that remain with a female to help guard and rear their young will have more and healthier offspring. Monogamy is observed in many bird populations where, in addition to the parental care from the female, the male is also a major provider of parental care for the chicks. A third explanation for the evolutionary advantages of monogamy is the “female-enforcement hypothesis.” In this scenario, the female ensures that the male does not have other offspring that might compete with her own, so she actively interferes with the male’s signaling to attract other mates.
Polygynous mating refers to one male mating with multiple females. In these situations, the female must be responsible for most of the parental care as the single male is not capable of providing care to that many offspring. In resourced-based polygyny, males compete for territories with the best resources, and then mate with females that enter the territory, drawn to its resource richness. The female benefits by mating with a dominant, genetically fit male; however, it is at the cost of having no male help in caring for the offspring. An example is seen in the yellow-rumped honeyguide, a bird whose males defend beehives because the females feed on their wax. As the females approach, the male defending the nest will mate with them. Harem mating structures are a type of polygynous system where certain males dominate mating while controlling a territory with resources. Elephant seals, where the alpha male dominates the mating within the group are an example. A third type of polygyny is a lek system. Here there is a communal courting area where several males perform elaborate displays for females, and the females choose their mate from this group. This behavior is observed in several bird species including the sage grouse and the prairie chicken.
In polyandrous mating systems, one female mates with many males. These types of systems are much rarer than monogamous and polygynous mating systems. In pipefishes and seahorses, males receive the eggs from the female, fertilize them, protect them within a pouch, and give birth to the offspring (Figure 23). Therefore, the female is able to provide eggs to several males without the burden of carrying the fertilized eggs.

Figure 23. Polyandrous mating, in which one female mates with many males, occurs in the (a) seahorse and the (b) pipefish. (credit a: modification of work by Brian Gratwicke; credit b: modification of work by Stephen Childs)
Learned Behaviors
Simple Learned Behaviors
The majority of the behaviors previously discussed were innate or at least have an innate component (variations on the innate behaviors may be learned). They are inherited and the behaviors do not change in response to signals from the environment. Conversely, learned behaviors, even though they may have instinctive components, allow an organism to adapt to changes in the environment and are modified by previous experiences. Simple learned behaviors include habituation and imprinting—both are important to the maturation process of young animals.
Habituation
Habituation is a simple form of learning in which an animal stops responding to a stimulus after a period of repeated exposure. This is a form of non-associative learning, as the stimulus is not associated with any punishment or reward. Prairie dogs typically sound an alarm call when threatened by a predator, but they become habituated to the sound of human footsteps when no harm is associated with this sound, therefore, they no longer respond to them with an alarm call. In this example, habituation is specific to the sound of human footsteps, as the animals still respond to the sounds of potential predators.
Imprinting

Figure 24. The attachment of ducklings to their mother is an example of imprinting. (credit: modification of work by Mark Harkin)
Imprinting is a type of learning that occurs at a particular age or a life stage that is rapid and independent of the species involved. Hatchling ducks recognize the first adult they see, their mother, and make a bond with her. A familiar sight is ducklings walking or swimming after their mothers (Figure 24). This is another type of non-associative learning, but is very important in the maturation process of these animals as it encourages them to stay near their mother so they will be protected, greatly increasing their chances of survival. However, if newborn ducks see a human before they see their mother, they will imprint on the human and follow it in just the same manner as they would follow their real mother.
The International Crane Foundation has helped raise the world’s population of whooping cranes from 21 individuals to about 600. Imprinting hatchlings has been a key to success: biologists wear full crane costumes so the birds never “see” humans. Watch this video to learn more.
Conditioned Behavior
Conditioned behaviors are types of associative learning, where a stimulus becomes associated with a consequence. During operant conditioning, the behavioral response is modified by its consequences, with regards to its form, strength, or frequency.
Classical Conditioning
In classical conditioning, a response called the conditioned response is associated with a stimulus that it had previously not been associated with, the conditioned stimulus. The response to the original, unconditioned stimulus is called the unconditioned response. The most cited example of classical conditioning is Ivan Pavlov’s experiments with dogs (Figure 25). In Pavlov’s experiments, the unconditioned response was the salivation of dogs in response to the unconditioned stimulus of seeing or smelling their food. The conditioning stimulus that researchers associated with the unconditioned response was the ringing of a bell. During conditioning, every time the animal was given food, the bell was rung. This was repeated during several trials. After some time, the dog learned to associate the ringing of the bell with food and to respond by salivating. After the conditioning period was finished, the dog would respond by salivating when the bell was rung, even when the unconditioned stimulus, the food, was absent. Thus, the ringing of the bell became the conditioned stimulus and the salivation became the conditioned response. Although it is thought by some scientists that the unconditioned and conditioned responses are identical, even Pavlov discovered that the saliva in the conditioned dogs had characteristic differences when compared to the unconditioned dog.
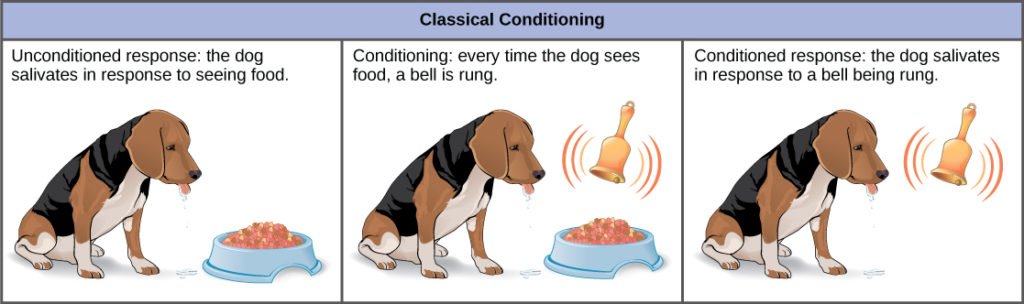
Figure 25. In the classic Pavlovian response, the dog becomes conditioned to associate the ringing of the bell with food.
It had been thought by some scientists that this type of conditioning required multiple exposures to the paired stimulus and response, but it is now known that this is not necessary in all cases, and that some conditioning can be learned in a single pairing experiment. Classical conditioning is a major tenet of behaviorism, a branch of psychological philosophy that proposes that all actions, thoughts, and emotions of living things are behaviors that can be treated by behavior modification and changes in the environment.
Operant Conditioning

Figure 26. The training of dolphins by rewarding them with food is an example of positive reinforcement operant conditioning. (credit: Roland Tanglao)
In operant conditioning, the conditioned behavior is gradually modified by its consequences as the animal responds to the stimulus. A major proponent of such conditioning was psychologist B.F. Skinner, the inventor of the Skinner box. Skinner put rats in his boxes that contained a lever that would dispense food to the rat when depressed. While initially the rat would push the lever a few times by accident, it eventually associated pushing the lever with getting the food. This type of learning is an example of operant conditioning. Operant learning is the basis of most animal training. The conditioned behavior is continually modified by positive or negative reinforcement, often a reward such as food or some type of punishment, respectively. In this way, the animal is conditioned to associate a type of behavior with the punishment or reward, and, over time, can be induced to perform behaviors that they would not have done in the wild, such as the “tricks” dolphins perform at marine amusement park shows (Figure 26).
Cognitive Learning
Classical and operant conditioning are inefficient ways for humans and other intelligent animals to learn. Some primates, including humans, are able to learn by imitating the behavior of others and by taking instructions. The development of complex language by humans has made cognitive learning, the manipulation of information using the mind, the most prominent method of human learning. In fact, that is how students are learning right now by reading this book. As students read, they can make mental images of objects or organisms and imagine changes to them, or behaviors by them, and anticipate the consequences. In addition to visual processing, cognitive learning is also enhanced by remembering past experiences, touching physical objects, hearing sounds, tasting food, and a variety of other sensory-based inputs. Cognitive learning is so powerful that it can be used to understand conditioning in detail. In the reverse scenario, conditioning cannot help someone learn about cognition.
Classic work on cognitive learning was done by Wolfgang Köhler with chimpanzees. He demonstrated that these animals were capable of abstract thought by showing that they could learn how to solve a puzzle. When a banana was hung in their cage too high for them to reach, and several boxes were placed randomly on the floor, some of the chimps were able to stack the boxes one on top of the other, climb on top of them, and get the banana. This implies that they could visualize the result of stacking the boxes even before they had performed the action. This type of learning is much more powerful and versatile than conditioning.
Cognitive learning is not limited to primates, although they are the most efficient in using it. Maze running experiments done with rats by H.C. Blodgett in the 1920s were the first to show cognitive skills in a simple mammal. The motivation for the animals to work their way through the maze was a piece of food at its end. In these studies, the animals in Group I were run in one trial per day and had food available to them each day on completion of the run (Figure 27). Group II rats were not fed in the maze for the first six days and then subsequent runs were done with food for several days after. Group III rats had food available on the third day and every day thereafter. The results were that the control rats, Group I, learned quickly, and figured out how to run the maze in seven days. Group III did not learn much during the three days without food, but rapidly caught up to the control group when given the food reward. Group II learned very slowly for the six days with no reward to motivate them, and they did not begin to catch up to the control group until the day food was given, and then it took two days longer to learn the maze.

Figure 27. Group I (the green solid line) found food at the end of each trial, group II (the blue dashed line) did not find food for the first 6 days, and group III (the red dotted line) did not find food during runs on the first three days. Notice that rats given food earlier learned faster and eventually caught up to the control group. The orange dots on the group II and III lines show the days when food rewards were added to the mazes.
It may not be immediately obvious that this type of learning is different than conditioning. Although one might be tempted to believe that the rats simply learned how to find their way through a conditioned series of right and left turns, E.C. Tolman proved a decade later that the rats were making a representation of the maze in their minds, which he called a “cognitive map.” This was an early demonstration of the power of cognitive learning and how these abilities were not just limited to humans.
Sociobiology
Sociobiology is an interdisciplinary science originally popularized by social insect researcher E.O. Wilson in the 1970s. Wilson defined the science as “the extension of population biology and evolutionary theory to social organization.”[1] The main thrust of sociobiology is that animal and human behavior, including aggressiveness and other social interactions, can be explained almost solely in terms of genetics and natural selection. This science is controversial; noted scientist such as the late Stephen Jay Gould criticized the approach for ignoring the environmental effects on behavior. This is another example of the “nature versus nurture” debate of the role of genetics versus the role of environment in determining an organism’s characteristics.
Sociobiology also links genes with behaviors and has been associated with “biological determinism,” the belief that all behaviors are hardwired into our genes. No one disputes that certain behaviors can be inherited and that natural selection plays a role retaining them. It is the application of such principles to human behavior that sparks this controversy, which remains active today.
Check Your Understanding
Answer the question(s) below to see how well you understand the topics covered in the previous section. This short quiz does not count toward your grade in the class, and you can retake it an unlimited number of times.
Use this quiz to check your understanding and decide whether to (1) study the previous section further or (2) move on to the next section.
Candela Citations
- Introduction to Community Ecology. Authored by: Shelli Carter and Lumen Learning. Provided by: Lumen Learning. License: CC BY: Attribution
- Biology. Provided by: OpenStax CNX. Located at: http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8. License: CC BY: Attribution. License Terms: Download for free at http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8
- Niches & competition. Provided by: Khan Academy. Located at: https://www.khanacademy.org/science/biology/ecology/community-ecosystem-ecology/a/niches-competition. License: CC BY-NC-SA: Attribution-NonCommercial-ShareAlike
- Anole Lizards. Authored by: Eva Horne. Provided by: OpenStax CNX. Located at: https://cnx.org/contents/24nI-KJ8@24.18:lGjgOeNc@8/Community-Ecology. Project: Principles of Biology. License: CC BY: Attribution
- Community Ecology: Feel the Love. Provided by: CrashCourse. Located at: https://youtu.be/GxE1SSqbSn4. Project: Crash Course Ecology. License: All Rights Reserved. License Terms: Standard YouTube License
- Edward O. Wilson. On Human Nature (1978; repr., Cambridge: Harvard University Press, 2004), xx. ↵