Identify the key elements and processes in plant growth
Most plants continue to grow throughout their lives. Like other multicellular organisms, plants grow through a combination of cell growth and cell division. Cell growth increases cell size, while cell division (mitosis) increases the number of cells.
Learning Objectives
- Describe the way that plants grow
- Distinguish between primary growth and secondary growth in stems
- Understand how hormones affect plant growth and development
How Plants Grow

Figure 1. There must be an area of growth, similar to how the bones in your fingers, arms, and legs grow longer. There is, and it is called the apical meristem, which is shown here.
Most plants continue to grow throughout their lives. Like other multicellular organisms, plants grow through a combination of cell growth and cell division. Cell growth increases cell size, while cell division (mitosis) increases the number of cells. As plant cells grow, they also become specialized into different cell types through cellular differentiation. Once cells differentiate, they can no longer divide. How do plants grow or replace damaged cells after that?
The key to continued growth and repair of plant cells is meristem. Meristem is a type of plant tissue consisting of undifferentiated cells that can continue to divide and differentiate.
Apical meristems are found at the apex, or tip, of roots and buds, allowing roots and stems to grow in length and leaves and flowers to differentiate. Roots and stems grow in length because the meristem adds tissue “behind” it, constantly propelling itself further into the ground (for roots) or air (for stems). Often, the apical meristem of a single branch will become dominant, suppressing the growth of meristems on other branches and leading to the development of a single trunk. In grasses, meristems at the base of the leaf blades allow for regrowth after grazing by herbivores—or mowing by lawnmowers.
Apical meristems differentiate into the three basic types of meristem tissue which correspond to the three types of tissue: protoderm produces new epidermis, ground meristem produces ground tissue, and procambium produces new xylem and phloem. These three types of meristem are considered primary meristem because they allow growth in length or height, which is known as primary growth.

Figure 2. Microphotograph of the root tip of a broad bean show rapidly dividing apical meristem tissue just behind the root cap. Numerous cells in various stages of mitosis can be observed.
Secondary meristems allow growth in diameter (secondary growth) in woody plants. Herbaceous plants do not have secondary growth. The two types of secondary meristem are both named cambium, meaning “exchange” or “change.” Vascular cambium produces secondary xylem (toward the center of the stem or root) and phloem (toward the outside of the stem or root), adding growth to the diameter of the plant. This process produces wood, and builds the sturdy trunks of trees. Cork cambium lies between the epidermis and the phloem, and replaces the epidermis of roots and stems with bark, one layer of which is cork.
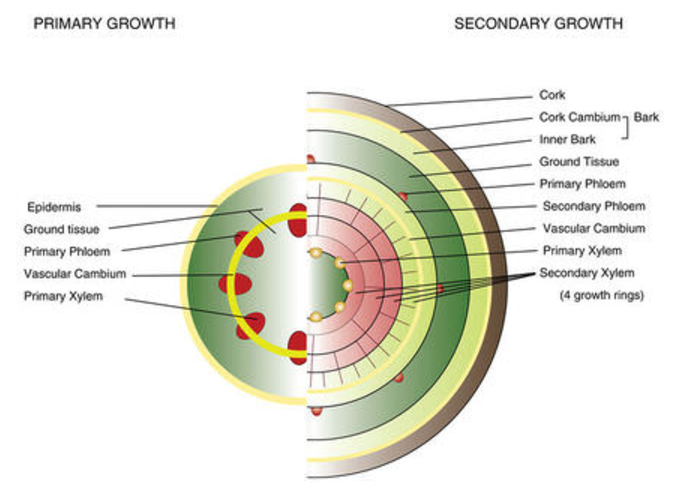
Figure 3. Primary and secondary growth
Woody plants grow in two ways. Primary growth adds length or height, mediated by apical meristem tissue at the tips of roots and shoots—which is difficult to show clearly in cross-sectional diagrams. Secondary growth adds to the diameter of a stem or root; vascular cambium adds xylem (inward) and phloem (outward), and cork cambium replaces epidermis with bark.
Watch this time-lapse video of plant growth. Note that there isn’t any narration in the video.
In Summary: How Plants Grow
Most plants continue to grow as long as they live. They grow through a combination of cell growth and cell division (mitosis). The key to plant growth is meristem, a type of plant tissue consisting of undifferentiated cells that can continue to divide and differentiate. Meristem allows plant stems and roots to grow longer (primary growth) and wider (secondary growth).
Stem Growth
Growth in plants occurs as the stems and roots lengthen. Some plants, especially those that are woody, also increase in thickness during their life span. The increase in length of the shoot and the root is referred to as primary growth, and is the result of cell division in the shoot apical meristem. Secondary growth is characterized by an increase in thickness or girth of the plant, and is caused by cell division in the lateral meristem. Figure 4 shows the areas of primary and secondary growth in a plant. Herbaceous plants mostly undergo primary growth, with hardly any secondary growth or increase in thickness. Secondary growth or “wood” is noticeable in woody plants; it occurs in some dicots, but occurs very rarely in monocots.

Figure 4. In woody plants, primary growth is followed by secondary growth, which allows the plant stem to increase in thickness or girth. Secondary vascular tissue is added as the plant grows, as well as a cork layer. The bark of a tree extends from the vascular cambium to the epidermis.
Some plant parts, such as stems and roots, continue to grow throughout a plant’s life: a phenomenon called indeterminate growth. Other plant parts, such as leaves and flowers, exhibit determinate growth, which ceases when a plant part reaches a particular size.
Primary Growth
Most primary growth occurs at the apices, or tips, of stems and roots. Primary growth is a result of rapidly dividing cells in the apical meristems at the shoot tip and root tip. Subsequent cell elongation also contributes to primary growth. The growth of shoots and roots during primary growth enables plants to continuously seek water (roots) or sunlight (shoots).
The influence of the apical bud on overall plant growth is known as apical dominance, which diminishes the growth of axillary buds that form along the sides of branches and stems. Most coniferous trees exhibit strong apical dominance, thus producing the typical conical Christmas tree shape. If the apical bud is removed, then the axillary buds will start forming lateral branches. Gardeners make use of this fact when they prune plants by cutting off the tops of branches, thus encouraging the axillary buds to grow out, giving the plant a bushy shape.
Secondary Growth
The increase in stem thickness that results from secondary growth is due to the activity of the lateral meristems, which are lacking in herbaceous plants. Lateral meristems include the vascular cambium and, in woody plants, the cork cambium (see Figure 4).

Figure 5. Lenticels on the bark of this cherry tree enable the woody stem to exchange gases with the surrounding atmosphere. (credit: Roger Griffith)
The vascular cambium is located just outside the primary xylem and to the interior of the primary phloem. The cells of the vascular cambium divide and form secondary xylem (tracheids and vessel elements) to the inside, and secondary phloem (sieve elements and companion cells) to the outside. The thickening of the stem that occurs in secondary growth is due to the formation of secondary phloem and secondary xylem by the vascular cambium, plus the action of cork cambium, which forms the tough outermost layer of the stem. The cells of the secondary xylem contain lignin, which provides hardiness and strength.
In woody plants, cork cambium is the outermost lateral meristem. It produces cork cells (bark) containing a waxy substance known as suberin that can repel water. The bark protects the plant against physical damage and helps reduce water loss. The cork cambium also produces a layer of cells known as phelloderm, which grows inward from the cambium. The cork cambium, cork cells, and phelloderm are collectively termed the periderm. The periderm substitutes for the epidermis in mature plants. In some plants, the periderm has many openings, known as lenticels, which allow the interior cells to exchange gases with the outside atmosphere (Figure 5). This supplies oxygen to the living and metabolically active cells of the cortex, xylem and phloem.
Annual Rings

Figure 6. The rate of wood growth increases in summer and decreases in winter, producing a characteristic ring for each year of growth. Seasonal changes in weather patterns can also affect the growth rate—note how the rings vary in thickness. (credit: Adrian Pingstone)
The activity of the vascular cambium gives rise to annual growth rings. During the spring growing season, cells of the secondary xylem have a large internal diameter and their primary cell walls are not extensively thickened. This is known as early wood, or spring wood. During the fall season, the secondary xylem develops thickened cell walls, forming late wood, or autumn wood, which is denser than early wood. This alternation of early and late wood is due largely to a seasonal decrease in the number of vessel elements and a seasonal increase in the number of tracheids. It results in the formation of an annual ring, which can be seen as a circular ring in the cross section of the stem (Figure 6). An examination of the number of annual rings and their nature (such as their size and cell wall thickness) can reveal the age of the tree and the prevailing climatic conditions during each season.
Growth Responses
A plant’s sensory response to external stimuli relies on chemical messengers (hormones). Plant hormones affect all aspects of plant life, from flowering to fruit setting and maturation, and from phototropism to leaf fall. Potentially every cell in a plant can produce plant hormones. They can act in their cell of origin or be transported to other portions of the plant body, with many plant responses involving the synergistic or antagonistic interaction of two or more hormones. In contrast, animal hormones are produced in specific glands and transported to a distant site for action, and they act alone.
Plant hormones are a group of unrelated chemical substances that affect plant morphogenesis. Five major plant hormones are traditionally described: auxins (particularly IAA), cytokinins, gibberellins, ethylene, and abscisic acid. In addition, other nutrients and environmental conditions can be characterized as growth factors.
Auxins
The term auxin is derived from the Greek word auxein, which means “to grow.” Auxins are the main hormones responsible for cell elongation in phototropism and gravitropism. They also control the differentiation of meristem into vascular tissue, and promote leaf development and arrangement. While many synthetic auxins are used as herbicides, IAA is the only naturally occurring auxin that shows physiological activity. Apical dominance—the inhibition of lateral bud formation—is triggered by auxins produced in the apical meristem. Flowering, fruit setting and ripening, and inhibition of abscission (leaf falling) are other plant responses under the direct or indirect control of auxins. Auxins also act as a relay for the effects of the blue light and red/far-red responses.
Commercial use of auxins is widespread in plant nurseries and for crop production. IAA is used as a rooting hormone to promote growth of adventitious roots on cuttings and detached leaves. Applying synthetic auxins to tomato plants in greenhouses promotes normal fruit development. Outdoor application of auxin promotes synchronization of fruit setting and dropping to coordinate the harvesting season. Fruits such as seedless cucumbers can be induced to set fruit by treating unfertilized plant flowers with auxins.
Cytokinins
The effect of cytokinins was first reported when it was found that adding the liquid endosperm of coconuts to developing plant embryos in culture stimulated their growth. The stimulating growth factor was found to be cytokinin, a hormone that promotes cytokinesis (cell division). Almost 200 naturally occurring or synthetic cytokinins are known to date. Cytokinins are most abundant in growing tissues, such as roots, embryos, and fruits, where cell division is occurring. Cytokinins are known to delay senescence in leaf tissues, promote mitosis, and stimulate differentiation of the meristem in shoots and roots. Many effects on plant development are under the influence of cytokinins, either in conjunction with auxin or another hormone. For example, apical dominance seems to result from a balance between auxins that inhibit lateral buds, and cytokinins that promote bushier growth.
Gibberellins

Figure 7. In grapes, application of gibberellic acid increases the size of fruit and loosens clustering. (credit: Bob Nichols, USDA)
Gibberellins (GAs) are a group of about 125 closely related plant hormones that stimulate shoot elongation, seed germination, and fruit and flower maturation. GAs are synthesized in the root and stem apical meristems, young leaves, and seed embryos. In urban areas, GA antagonists are sometimes applied to trees under power lines to control growth and reduce the frequency of pruning.
GAs break dormancy (a state of inhibited growth and development) in the seeds of plants that require exposure to cold or light to germinate. Abscisic acid is a strong antagonist of GA action. Other effects of GAs include gender expression, seedless fruit development, and the delay of senescence in leaves and fruit. Seedless grapes are obtained through standard breeding methods and contain inconspicuous seeds that fail to develop. Because GAs are produced by the seeds, and because fruit development and stem elongation are under GA control, these varieties of grapes would normally produce small fruit in compact clusters. Maturing grapes are routinely treated with GA to promote larger fruit size, as well as looser bunches (longer stems), which reduces the instance of mildew infection (Figure 7).
Abscisic Acid
The plant hormone abscisic acid (ABA) was first discovered as the agent that causes the abscission or dropping of cotton bolls. However, more recent studies indicate that ABA plays only a minor role in the abscission process. ABA accumulates as a response to stressful environmental conditions, such as dehydration, cold temperatures, or shortened day lengths. Its activity counters many of the growth-promoting effects of GAs and auxins. ABA inhibits stem elongation and induces dormancy in lateral buds.
ABA induces dormancy in seeds by blocking germination and promoting the synthesis of storage proteins. Plants adapted to temperate climates require a long period of cold temperature before seeds germinate. This mechanism protects young plants from sprouting too early during unseasonably warm weather in winter. As the hormone gradually breaks down over winter, the seed is released from dormancy and germinates when conditions are favorable in spring. Another effect of ABA is to promote the development of winter buds; it mediates the conversion of the apical meristem into a dormant bud. Low soil moisture causes an increase in ABA, which causes stomata to close, reducing water loss in winter buds.
Ethylene
Ethylene is associated with fruit ripening, flower wilting, and leaf fall. Ethylene is unusual because it is a volatile gas (C2H4). Hundreds of years ago, when gas street lamps were installed in city streets, trees that grew close to lamp posts developed twisted, thickened trunks and shed their leaves earlier than expected. These effects were caused by ethylene volatilizing from the lamps.
Aging tissues (especially senescing leaves) and nodes of stems produce ethylene. The best-known effect of the hormone, however, is the promotion of fruit ripening. Ethylene stimulates the conversion of starch and acids to sugars. Some people store unripe fruit, such as avocadoes, in a sealed paper bag to accelerate ripening; the gas released by the first fruit to mature will speed up the maturation of the remaining fruit. Ethylene also triggers leaf and fruit abscission, flower fading and dropping, and promotes germination in some cereals and sprouting of bulbs and potatoes.
Ethylene is widely used in agriculture. Commercial fruit growers control the timing of fruit ripening with application of the gas. Horticulturalists inhibit leaf dropping in ornamental plants by removing ethylene from greenhouses using fans and ventilation.
Nontraditional Hormones
Recent research has discovered a number of compounds that also influence plant development. Their roles are less understood than the effects of the major hormones described so far.
Jasmonates play a major role in defense responses to herbivory. Their levels increase when a plant is wounded by a predator, resulting in an increase in toxic secondary metabolites. They contribute to the production of volatile compounds that attract natural enemies of predators. For example, chewing of tomato plants by caterpillars leads to an increase in jasmonic acid levels, which in turn triggers the release of volatile compounds that attract predators of the pest.
Oligosaccharins also play a role in plant defense against bacterial and fungal infections. They act locally at the site of injury, and can also be transported to other tissues. Strigolactones promote seed germination in some species and inhibit lateral apical development in the absence of auxins. Strigolactones also play a role in the establishment of mycorrhizae, a mutualistic association of plant roots and fungi. Brassinosteroids are important to many developmental and physiological processes. Signals between these compounds and other hormones, notably auxin and GAs, amplifies their physiological effect. Apical dominance, seed germination, gravitropism, and resistance to freezing are all positively influenced by hormones. Root growth and fruit dropping are inhibited by steroids.
Check Your Understanding
Answer the question(s) below to see how well you understand the topics covered in the previous section. This short quiz does not count toward your grade in the class, and you can retake it an unlimited number of times.
Use this quiz to check your understanding and decide whether to (1) study the previous section further or (2) move on to the next section.
Candela Citations
- Introduction to Plant Growth. Authored by: Shelli Carter and Lumen Learning. Provided by: Lumen Learning. License: CC BY: Attribution
- Plant Growth. Authored by: CK-12. Located at: http://www.ck12.org/biology/Plant-Growth/lesson/Plant-Growth/r35/. Project: Biology. License: CC BY-NC: Attribution-NonCommercial
- Stem Growth and Growth Responses. Provided by: OpenStax. Located at: http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8. Project: Biology. License: CC BY: Attribution. License Terms: Download for free at http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8