Classify seed plants
The lush palms on tropical shorelines do not depend on water for the dispersal of their pollen, fertilization, or the survival of the zygote—unlike mosses, liverworts, and ferns of the terrain. Seed plants, such as palms, have broken free from the need to rely on water for their reproductive needs. They play an integral role in all aspects of life on the planet, shaping the physical terrain, influencing the climate, and maintaining life as we know it. For millennia, human societies have depended on seed plants for nutrition and medicinal compounds: and more recently, for industrial by-products, such as timber and paper, dyes, and textiles. Palms provide materials including rattans, oils, and dates. Wheat is grown to feed both human and animal populations. The fruit of the cotton boll flower is harvested as a boll, with its fibers transformed into clothing or pulp for paper. The showy opium poppy is valued both as an ornamental flower and as a source of potent opiate compounds.

Figure 1. Seed plants dominate the landscape and play an integral role in human societies. (a) Palm trees grow along the shoreline; (b) wheat is a crop grown in most of the world; (c) the flower of the cotton plant produces fibers that are woven into fabric; (d) the potent alkaloids of the beautiful opium poppy have influenced human life both as a medicinal remedy and as a dangerously addictive drug. (credit a: modification of work by Ryan Kozie; credit b: modification of work by Stephen Ausmus; credit c: modification of work by David Nance; credit d: modification of work by Jolly Janner)
Learning Objectives
- Describe the evolutionary history of seed plants
- Identify the main characteristics of gymnosperms
- Identify the main characteristics of angiosperms
- Discuss the roles that plants play in ecosystems
Evolution of Seed Plants
The first plants to colonize land were most likely closely related to modern day mosses (bryophytes) and are thought to have appeared about 500 million years ago. They were followed by liverworts (also bryophytes) and primitive vascular plants—the pterophytes—from which modern ferns are derived. The lifecycle of bryophytes and pterophytes is characterized by the alternation of generations, like gymnosperms and angiosperms; what sets bryophytes and pterophytes apart from gymnosperms and angiosperms is their reproductive requirement for water. The completion of the bryophyte and pterophyte life cycle requires water because the male gametophyte releases sperm, which must swim—propelled by their flagella—to reach and fertilize the female gamete or egg. After fertilization, the zygote matures and grows into a sporophyte, which in turn will form sporangia or “spore vessels.” In the sporangia, mother cells undergo meiosis and produce the haploid spores. Release of spores in a suitable environment will lead to germination and a new generation of gametophytes.
In seed plants, the evolutionary trend led to a dominant sporophyte generation, and at the same time, a systematic reduction in the size of the gametophyte: from a conspicuous structure to a microscopic cluster of cells enclosed in the tissues of the sporophyte. Whereas lower vascular plants, such as club mosses and ferns, are mostly homosporous (produce only one type of spore), all seed plants, or spermatophytes, are heterosporous. They form two types of spores: megaspores (female) and microspores (male). Megaspores develop into female gametophytes that produce eggs, and microspores mature into male gametophytes that generate sperm. Because the gametophytes mature within the spores, they are not free-living, as are the gametophytes of other seedless vascular plants. Heterosporous seedless plants are seen as the evolutionary forerunners of seed plants.
Seeds and pollen—two critical adaptations to drought, and to reproduction that doesn’t require water—distinguish seed plants from other (seedless) vascular plants. Both adaptations were required for the colonization of land begun by the bryophytes and their ancestors. Fossils place the earliest distinct seed plants at about 350 million years ago. The first reliable record of gymnosperms dates their appearance to the Pennsylvanian period, about 319 million years ago (Table 1). Gymnosperms were preceded by progymnosperms, the first naked seed plants, which arose about 380 million years ago. Progymnosperms were a transitional group of plants that superficially resembled conifers (cone bearers) because they produced wood from the secondary growth of the vascular tissues; however, they still reproduced like ferns, releasing spores into the environment. Gymnosperms dominated the landscape in the early (Triassic) and middle (Jurassic) Mesozoic era. Angiosperms surpassed gymnosperms by the middle of the Cretaceous (about 100 million years ago) in the late Mesozoic era, and today are the most abundant plant group in most terrestrial biomes.
| Table 1. Geologic Timescale[1] | |||
|---|---|---|---|
| Eon | Era | Period | Start, MYA |
| Phanerozoic | Cenozoic | Quaternary | 1.6 |
| Neogene | 23 | ||
| Paleogene | 66 | ||
| Mesozoic | Cretaceous | 145 | |
| Jurassic | 201 | ||
| Triassic | 252 | ||
| Paleozoic | Permian | 298 | |
| Carboniferous | 360 | ||
| Devonian | 419 | ||
| Silurian | 444 | ||
| Ordovician | 485 | ||
| Cambrian | 540 | ||
| Proterozoic | Late Proterozoic Middle Proterozoic Early Proterozoic |
2500 | |
| Archean | Late Archean Middle Archean Early Archean |
4000 | |
| Pre-Archean | ~4600 | ||
Pollen and seed were innovative structures that allowed seed plants to break their dependence on water for reproduction and development of the embryo, and to conquer dry land. The pollen grains are the male gametophytes, which contain the sperm (gametes) of the plant. The small haploid (1n) cells are encased in a protective coat that prevents desiccation (drying out) and mechanical damage. Pollen grains can travel far from their original sporophyte, spreading the plant’s genes. The seed offers the embryo protection, nourishment, and a mechanism to maintain dormancy for tens or even thousands of years, ensuring germination can occur when growth conditions are optimal. Seeds therefore allow plants to disperse the next generation through both space and time. With such evolutionary advantages, seed plants have become the most successful and familiar group of plants, in part because of their size and striking appearance.
Evolution of Gymnosperms

Figure 2. This fossilized leaf is from Glossopteris, a seed fern that thrived during the Permian age (290–240 million years ago). (credit: D.L. Schmidt, USGS)
The fossil plant Elkinsia polymorpha, a “seed fern” from the Devonian period—about 400 million years ago—is considered the earliest seed plant known to date. Seed ferns (Figure 2) produced their seeds along their branches without specialized structures. What makes them the first true seed plants is that they developed structures called cupules to enclose and protect the ovule—the female gametophyte and associated tissues—which develops into a seed upon fertilization. Seed plants resembling modern tree ferns became more numerous and diverse in the coal swamps of the Carboniferous period.
Fossil records indicate the first gymnosperms (progymnosperms) most likely originated in the Paleozoic era, during the middle Devonian period: about 390 million years ago. Following the wet Mississippian and Pennsylvanian periods, which were dominated by giant fern trees, the Permian period was dry. This gave a reproductive edge to seed plants, which are better adapted to survive dry spells.

Figure 3. This boreal forest (taiga) has low-lying plants and conifer trees. (credit: L.B. Brubaker, NOAA)
The Ginkgoales, a group of gymnosperms with only one surviving species—the Gingko biloba—were the first gymnosperms to appear during the lower Jurassic. Gymnosperms expanded in the Mesozoic era (about 240 million years ago), supplanting ferns in the landscape, and reaching their greatest diversity during this time. The Jurassic period was as much the age of the cycads (palm-tree-like gymnosperms) as the age of the dinosaurs. Gingkoales and the more familiar conifers also dotted the landscape. Although angiosperms (flowering plants) are the major form of plant life in most biomes, gymnosperms still dominate some ecosystems, such as the taiga (boreal forests) and the alpine forests at higher mountain elevations (Figure 3) because of their adaptation to cold and dry growth conditions.
Seeds and Pollen as an Evolutionary Adaptation to Dry Land

Figure 4. This fossilized pollen is from a Buckbean fen core found in Yellowstone National Park, Wyoming. The pollen is magnified 1,054 times. (credit: R.G. Baker, USGS; scale-bar data from Matt Russell)
Unlike bryophyte and fern spores (which are haploid cells dependent on moisture for rapid development of gametophytes), seeds contain a diploid embryo that will germinate into a sporophyte. Storage tissue to sustain growth and a protective coat give seeds their superior evolutionary advantage. Several layers of hardened tissue prevent desiccation, and free reproduction from the need for a constant supply of water. Furthermore, seeds remain in a state of dormancy—induced by desiccation and the hormone abscisic acid—until conditions for growth become favorable. Whether blown by the wind, floating on water, or carried away by animals, seeds are scattered in an expanding geographic range, thus avoiding competition with the parent plant.
Pollen grains (Figure 4) are male gametophytes and are carried by wind, water, or a pollinator. The whole structure is protected from desiccation and can reach the female organs without dependence on water. Male gametes reach female gametophyte and the egg cell gamete though a pollen tube: an extension of a cell within the pollen grain. The sperm of modern gymnosperms lack flagella, but in cycads and the Gingko, the sperm still possess flagella that allow them to swim down the pollen tube to the female gamete; however, they are enclosed in a pollen grain.
Evolution of Angiosperms

Figure 5. This leaf imprint shows a Ficus speciosissima, an angiosperm that flourished during the Cretaceous period. (credit: W. T. Lee, USGS)
Undisputed fossil records place the massive appearance and diversification of angiosperms in the middle to late Mesozoic era. Angiosperms (“seed in a vessel”) produce a flower containing male and/or female reproductive structures. Fossil evidence (Figure 5) indicates that flowering plants first appeared in the Lower Cretaceous, about 125 million years ago, and were rapidly diversifying by the Middle Cretaceous, about 100 million years ago.
Earlier traces of angiosperms are scarce. Fossilized pollen recovered from Jurassic geological material has been attributed to angiosperms. A few early Cretaceous rocks show clear imprints of leaves resembling angiosperm leaves. By the mid-Cretaceous, a staggering number of diverse flowering plants crowd the fossil record. The same geological period is also marked by the appearance of many modern groups of insects, including pollinating insects that played a key role in ecology and the evolution of flowering plants.
Although several hypotheses have been offered to explain this sudden profusion and variety of flowering plants, none have garnered the consensus of paleobotanists (scientists who study ancient plants). New data in comparative genomics and paleobotany have, however, shed some light on the evolution of angiosperms. Rather than being derived from gymnosperms, angiosperms form a sister clade (a species and its descendents) that developed in parallel with the gymnosperms. The two innovative structures of flowers and fruit represent an improved reproductive strategy that served to protect the embryo, while increasing genetic variability and range. Paleobotanists debate whether angiosperms evolved from small woody bushes, or were basal angiosperms related to tropical grasses. Both views draw support from cladistics studies, and the so-called woody magnoliid hypothesis—which proposes that the early ancestors of angiosperms were shrubs—also offers molecular biological evidence.
The most primitive living angiosperm is considered to be Amborella trichopoda, a small plant native to the rainforest of New Caledonia, an island in the South Pacific. Analysis of the genome of A. trichopoda has shown that it is related to all existing flowering plants and belongs to the oldest confirmed branch of the angiosperm family tree. A few other angiosperm groups called basal angiosperms, are viewed as primitive because they branched off early from the phylogenetic tree. Most modern angiosperms are classified as either monocots or eudicots, based on the structure of their leaves and embryos. Basal angiosperms, such as water lilies, are considered more primitive because they share morphological traits with both monocots and eudicots.
Flowers and Fruits as an Evolutionary Adaptation
Angiosperms produce their gametes in separate organs, which are usually housed in a flower. Both fertilization and embryo development take place inside an anatomical structure that provides a stable system of sexual reproduction largely sheltered from environmental fluctuations. Flowering plants are the most diverse phylum on Earth after insects; flowers come in a bewildering array of sizes, shapes, colors, smells, and arrangements. Most flowers have a mutualistic pollinator, with the distinctive features of flowers reflecting the nature of the pollination agent. The relationship between pollinator and flower characteristics is one of the great examples of coevolution.
Following fertilization of the egg, the ovule grows into a seed. The surrounding tissues of the ovary thicken, developing into a fruit that will protect the seed and often ensure its dispersal over a wide geographic range. Not all fruits develop from an ovary; such structures are “false fruits.” Like flowers, fruit can vary tremendously in appearance, size, smell, and taste. Tomatoes, walnut shells and avocados are all examples of fruit. As with pollen and seeds, fruits also act as agents of dispersal. Some may be carried away by the wind. Many attract animals that will eat the fruit and pass the seeds through their digestive systems, then deposit the seeds in another location. Cockleburs are covered with stiff, hooked spines that can hook into fur (or clothing) and hitch a ride on an animal for long distances. The cockleburs that clung to the velvet trousers of an enterprising Swiss hiker, George de Mestral, inspired his invention of the loop and hook fastener he named Velcro.
Building Phylogenetic Trees with Analysis of DNA Sequence Alignments
All living organisms display patterns of relationships derived from their evolutionary history. Phylogeny is the science that describes the relative connections between organisms, in terms of ancestral and descendant species. Phylogenetic trees, such as the plant evolutionary history shown in Figure 6, are tree-like branching diagrams that depict these relationships. Species are found at the tips of the branches. Each branching point, called a node, is the point at which a single taxonomic group (taxon), such as a species, separates into two or more species.

Figure 6. This phylogenetic tree shows the evolutionary relationships of plants.
Phylogenetic trees have been built to describe the relationships between species since Darwin’s time. Traditional methods involve comparison of homologous anatomical structures and embryonic development, assuming that closely related organisms share anatomical features during embryo development. Some traits that disappear in the adult are present in the embryo; for example, a human fetus, at one point, has a tail. The study of fossil records shows the intermediate stages that link an ancestral form to its descendants. Most of these approaches are imprecise and lend themselves to multiple interpretations. As the tools of molecular biology and computational analysis have been developed and perfected in recent years, a new generation of tree-building methods has taken shape. The key assumption is that genes for essential proteins or RNA structures, such as the ribosomal RNA, are inherently conserved because mutations (changes in the DNA sequence) could compromise the survival of the organism. DNA from minute amounts of living organisms or fossils can be amplified by polymerase chain reaction (PCR) and sequenced, targeting the regions of the genome that are most likely to be conserved between species. The genes encoding the ribosomal RNA from the small 18S subunit and plastid genes are frequently chosen for DNA alignment analysis.
Once the sequences of interest are obtained, they are compared with existing sequences in databases such as GenBank, which is maintained by The National Center for Biotechnology Information. A number of computational tools are available to align and analyze sequences. Sophisticated computer analysis programs determine the percentage of sequence identity or homology. Sequence homology can be used to estimate the evolutionary distance between two DNA sequences and reflect the time elapsed since the genes separated from a common ancestor. Molecular analysis has revolutionized phylogenetic trees. In some cases, prior results from morphological studies have been confirmed: for example, confirming Amborella trichopoda as the most primitive angiosperm known. However, some groups and relationships have been rearranged as a result of DNA analysis.
Gymnosperms
Gymnosperms, meaning “naked seeds,” are a diverse group of seed plants and are paraphyletic. Paraphyletic groups are those in which not all members are descendants of a single common ancestor. Their characteristics include naked seeds, separate female and male gametes, pollination by wind, and tracheids (which transport water and solutes in the vascular system).
Gymnosperm seeds are not enclosed in an ovary; rather, they are exposed on cones or modified leaves. Sporophylls are specialized leaves that produce sporangia. The term strobilus (plural = strobili) describes a tight arrangement of sporophylls around a central stalk, as seen in cones. Some seeds are enveloped by sporophyte tissues upon maturation. The layer of sporophyte tissue that surrounds the megasporangium, and later, the embryo, is called the integument.
Gymnosperms were the dominant phylum in Mesozoic era. They are adapted to live where fresh water is scarce during part of the year, or in the nitrogen-poor soil of a bog. Therefore, they are still the prominent phylum in the coniferous biome or taiga, where the evergreen conifers have a selective advantage in cold and dry weather. Evergreen conifers continue low levels of photosynthesis during the cold months, and are ready to take advantage of the first sunny days of spring. One disadvantage is that conifers are more susceptible than deciduous trees to infestations because conifers do not lose their leaves all at once. They cannot, therefore, shed parasites and restart with a fresh supply of leaves in spring.
The life cycle of a gymnosperm involves alternation of generations, with a dominant sporophyte in which the female gametophyte resides, and reduced gametophytes. All gymnosperms are heterosporous. The male and female reproductive organs can form in cones or strobili. Male and female sporangia are produced either on the same plant, described as monoecious (“one home” or bisexual), or on separate plants, referred to as dioecious (“two homes” or unisexual) plants. The life cycle of a conifer will serve as our example of reproduction in gymnosperms.
Life Cycle of a Conifer
Pine trees are conifers (cone bearing) and carry both male and female sporophylls on the same mature sporophyte. Therefore, they are monoecious plants. Like all gymnosperms, pines are heterosporous and generate two different types of spores: male microspores and female megaspores. In the male cones, or staminate cones, the microsporocytes give rise to pollen grains by meiosis. In the spring, large amounts of yellow pollen are released and carried by the wind. Some gametophytes will land on a female cone. Pollination is defined as the initiation of pollen tube growth. The pollen tube develops slowly, and the generative cell in the pollen grain divides into two haploid sperm cells by mitosis. At fertilization, one of the sperm cells will finally unite its haploid nucleus with the haploid nucleus of a haploid egg cell.
Female cones, or ovulate cones, contain two ovules per scale. One megaspore mother cell, or megasporocyte, undergoes meiosis in each ovule. Three of the four cells break down; only a single surviving cell will develop into a female multicellular gametophyte, which encloses archegonia (an archegonium is a reproductive organ that contains a single large egg). Upon fertilization, the diploid egg will give rise to the embryo, which is enclosed in a seed coat of tissue from the parent plant. Fertilization and seed development is a long process in pine trees: it may take up to two years after pollination. The seed that is formed contains three generations of tissues: the seed coat that originates from the sporophyte tissue, the gametophyte that will provide nutrients, and the embryo itself.
Figure 7 illustrates the life cycle of a conifer. The sporophyte (2n) phase is the longest phase in the life of a gymnosperm. The gametophytes (1n)—microspores and megaspores—are reduced in size. It may take more than year between pollination and fertilization while the pollen tube grows towards the megasporocyte (2n), which undergoes meiosis into megaspores. The megaspores will mature into eggs (1n).

Figure 7. This image shows the life cycle of a conifer. Pollen from male cones blows up into upper branches, where it fertilizes female cones.
Practice Question
At what stage does the diploid zygote form?
- when the female cone begins to bud from the tree
- at fertilization
- when the seeds drop from the tree
- when the pollen tube begins to grow
Watch this video to see the process of seed production in gymnosperms.
Diversity of Gymnosperms
Modern gymnosperms are classified into four phyla. Coniferophyta, Cycadophyta, and Ginkgophyta are similar in their production of secondary cambium (cells that generate the vascular system of the trunk or stem and are partially specialized for water transportation) and their pattern of seed development. However, the three phyla are not closely related phylogenetically to each other. Gnetophyta are considered the closest group to angiosperms because they produce true xylem tissue.
Conifers (Coniferophyta)
Conifers are the dominant phylum of gymnosperms, with the most variety of species (Figure 8). Most are typically tall trees that usually bear scale-like or needle-like leaves. Water evaporation from leaves is reduced by their thin shape and the thick cuticle. Snow slides easily off needle-shaped leaves, keeping the load light and decreasing breaking of branches. Adaptations to cold and dry weather explain the predominance of conifers at high altitudes and in cold climates. Conifers include familiar evergreen trees such as pines, spruces, firs, cedars, sequoias, and yews. A few species are deciduous and lose their leaves in fall. The European larch and the tamarack are examples of deciduous conifers (Figure 8c). Many coniferous trees are harvested for paper pulp and timber. The wood of conifers is more primitive than the wood of angiosperms; it contains tracheids, but no vessel elements, and is therefore referred to as “soft wood.”

Figure 8. Conifers are the dominant form of vegetation in cold or arid environments and at high altitudes. Shown here are the (a) evergreen spruce Picea sp., (b) juniper Juniperus sp., (c) sequoia Sequoia Semervirens, which is a deciduous gymnosperm, and (d) the tamarack Larix larcinia. Notice the yellow leaves of the tamarack. (credit a: modification of work by Rosendahl; credit b: modification of work by Alan Levine; credit c: modification of work by Wendy McCormic; credit d: modification of work by Micky Zlimen)
Cycads

Figure 9. This Encephalartos ferox cycad has large cones and broad, fern-like leaves. (credit: Wendy Cutler)
Cycads thrive in mild climates, and are often mistaken for palms because of the shape of their large, compound leaves. Cycads bear large cones (Figure 9), and may be pollinated by beetles rather than wind: unusual for a gymnosperm. They dominated the landscape during the age of dinosaurs in the Mesozoic, but only a hundred or so species persisted to modern times. They face possible extinction, and several species are protected through international conventions. Because of their attractive shape, they are often used as ornamental plants in gardens in the tropics and subtropics.
Gingkophytes
The single surviving species of the gingkophytes group is the Gingko biloba (Figure 10). Its fan-shaped leaves—unique among seed plants because they feature a dichotomous venation pattern—turn yellow in autumn and fall from the tree. For centuries, G. bilobawas cultivated by Chinese Buddhist monks in monasteries, which ensured its preservation. It is planted in public spaces because it is unusually resistant to pollution. Male and female organs are produced on separate plants. Typically, gardeners plant only male trees because the seeds produced by the female plant have an off-putting smell of rancid butter.

Figure 10. This plate from the 1870 book Flora Japonica, Sectio Prima (Tafelband) depicts the leaves and fruit of Gingko biloba, as drawn by Philipp Franz von Siebold and Joseph Gerhard Zuccarini.
Gnetophytes
Gnetophytes are the closest relative to modern angiosperms, and include three dissimilar genera of plants: Ephedra, Gnetum, and Welwitschia (Figure 11). Like angiosperms, they have broad leaves. In tropical and subtropical zones, gnetophytes are vines or small shrubs. Ephedra occurs in dry areas of the West Coast of the United States and Mexico. Ephedra’s small, scale-like leaves are the source of the compound ephedrine, which is used in medicine as a potent decongestant. Because ephedrine is similar to amphetamines, both in chemical structure and neurological effects, its use is restricted to prescription drugs. Like angiosperms, but unlike other gymnosperms, all gnetophytes possess vessel elements in their xylem.

Figure 11. (a) Ephedra viridis, known by the common name Mormon tea, grows on the West Coast of the United States and Mexico. (b)Gnetum gnemon grows in Malaysia. (c) The large Welwitschia mirabilis can be found in the Namibian desert. (credit a: modification of work by USDA; credit b: modification of work by Malcolm Manners; credit c: modification of work by Derek Keats)
Watch this video describing the amazing strangeness of Welwitschia.
Angiosperms

Figure 12. These flowers grow in a botanical garden border in Bellevue, WA. Flowering plants dominate terrestrial landscapes. The vivid colors of flowers are an adaptation to pollination by animals such as insects and birds. (credit: Myriam Feldman)
From their humble and still obscure beginning during the early Jurassic period, the angiosperms—or flowering plants—have evolved to dominate most terrestrial ecosystems (Figure 12). With more than 250,000 species, the angiosperm phylum (Anthophyta) is second only to insects in terms of diversification.
The success of angiosperms is due to two novel reproductive structures: flowers and fruit. The function of the flower is to ensure pollination. Flowers also provide protection for the ovule and developing embryo inside a receptacle. The function of the fruit is seed dispersal. They also protect the developing seed. Different fruit structures or tissues on fruit—such as sweet flesh, wings, parachutes, or spines that grab—reflect the dispersal strategies that help spread seeds.
Flowers
Flowers are modified leaves, or sporophylls, organized around a central stalk. Although they vary greatly in appearance, all flowers contain the same structures: sepals, petals, carpels, and stamens. The peduncle attaches the flower to the plant. A whorl of sepals (collectively called the calyx) is located at the base of the peduncle and encloses the unopened floral bud. Sepals are usually photosynthetic organs, although there are some exceptions. For example, the corolla in lilies and tulips consists of three sepals and three petals that look virtually identical. Petals, collectively the corolla, are located inside the whorl of sepals and often display vivid colors to attract pollinators. Flowers pollinated by wind are usually small, feathery, and visually inconspicuous. Sepals and petals together form the perianth. The sexual organs (carpels and stamens) are located at the center of the flower.
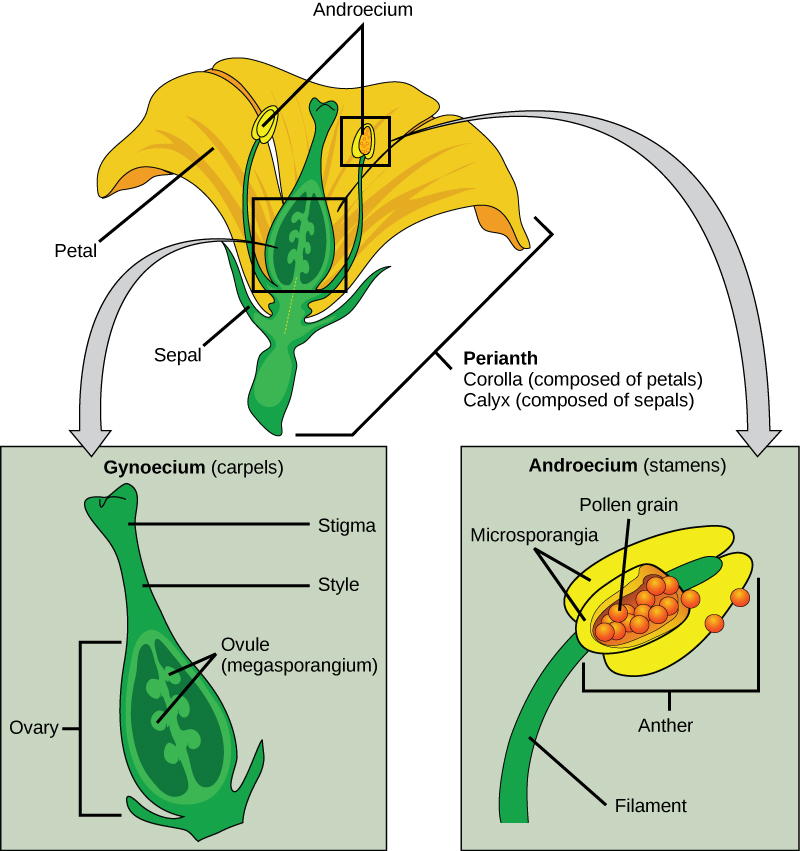
Figure 13. This image depicts the structure of a perfect flower. Perfect flowers produce both male and female floral organs. The flower shown has only one carpel, but some flowers have a cluster of carpels. Together, all the carpels make up the gynoecium. (credit: modification of work by Mariana Ruiz Villareal)
As illustrated in Figure 13, styles, stigmas, and ovules constitute the female organ: the gynoecium or carpel. Flower structure is very diverse, and carpels may be singular, multiple, or fused. Multiple fused carpels comprise a pistil. The megaspores and the female gametophytes are produced and protected by the thick tissues of the carpel. A long, thin structure called a style leads from the sticky stigma, where pollen is deposited, to the ovary, enclosed in the carpel. The ovary houses one or more ovules, each of which will develop into a seed upon fertilization. The male reproductive organs, the stamens (collectively called the androecium), surround the central carpel. Stamens are composed of a thin stalk called a filament and a sac-like structure called the anther. The filament supports the anther, where the microspores are produced by meiosis and develop into pollen grains.
Fruit
As the seed develops, the walls of the ovary thicken and form the fruit. The seed forms in an ovary, which also enlarges as the seeds grow. In botany, a fertilized and fully grown, ripened ovary is a fruit. Many foods commonly called vegetables are actually fruit. Eggplants, zucchini, string beans, and bell peppers are all technically fruit because they contain seeds and are derived from the thick ovary tissue. Acorns are nuts, and winged maple whirligigs (whose botanical name is samara) are also fruit. Botanists classify fruit into more than two dozen different categories, only a few of which are actually fleshy and sweet.
Mature fruit can be fleshy or dry. Fleshy fruit include the familiar berries, peaches, apples, grapes, and tomatoes. Rice, wheat, and nuts are examples of dry fruit. Another distinction is that not all fruits are derived from the ovary. For instance, strawberries are derived from the receptacle and apples from the pericarp, or hypanthium. Some fruits are derived from separate ovaries in a single flower, such as the raspberry. Other fruits, such as the pineapple, form from clusters of flowers. Additionally, some fruits, like watermelon and orange, have rinds. Regardless of how they are formed, fruits are an agent of seed dispersal. The variety of shapes and characteristics reflect the mode of dispersal. Wind carries the light dry fruit of trees and dandelions. Water transports floating coconuts. Some fruits attract herbivores with color or perfume, or as food. Once eaten, tough, undigested seeds are dispersed through the herbivore’s feces. Other fruits have burs and hooks to cling to fur and hitch rides on animals.
The Life Cycle of an Angiosperm
The adult, or sporophyte, phase is the main phase of an angiosperm’s life cycle (Figure 14). Like gymnosperms, angiosperms are heterosporous. Therefore, they generate microspores, which will generate pollen grains as the male gametophytes, and megaspores, which will form an ovule that contains female gametophytes. Inside the anthers’ microsporangia, male gametophytes divide by meiosis to generate haploid microspores, which, in turn, undergo mitosis and give rise to pollen grains. Each pollen grain contains two cells: one generative cell that will divide into two sperm and a second cell that will become the pollen tube cell.

Figure 14. The life cycle of an angiosperm is shown. Anthers and carpels are structures that shelter the actual gametophytes: the pollen grain and embryo sac. Double fertilization is a process unique to angiosperms. (credit: modification of work by Mariana Ruiz Villareal)
Practice Question
If a flower lacked a megasporangium, what type of gamete would not form?
If the flower lacked a microsporangium, what type of gamete would not form?
The ovule, sheltered within the ovary of the carpel, contains the megasporangium protected by two layers of integuments and the ovary wall. Within each megasporangium, a megasporocyte undergoes meiosis, generating four megaspores—three small and one large. Only the large megaspore survives; it produces the female gametophyte, referred to as the embryo sac. The megaspore divides three times to form an eight-cell stage. Four of these cells migrate to each pole of the embryo sac; two come to the equator, and will eventually fuse to form a 2n polar nucleus; the three cells away from the egg form antipodals, and the two cells closest to the egg become the synergids.
The mature embryo sac contains one egg cell, two synergids or “helper” cells, three antipodal cells, and two polar nuclei in a central cell. When a pollen grain reaches the stigma, a pollen tube extends from the grain, grows down the style, and enters through the micropyle: an opening in the integuments of the ovule. The two sperm cells are deposited in the embryo sac.
A double fertilization event then occurs. One sperm and the egg combine, forming a diploid zygote—the future embryo. The other sperm fuses with the 2n polar nuclei, forming a triploid cell that will develop into the endosperm, which is tissue that serves as a food reserve. The zygote develops into an embryo with a radicle, or small root, and one (monocot) or two (dicot) leaf-like organs called cotyledons. This difference in the number of embryonic leaves is the basis for the two major groups of angiosperms: the monocots and the eudicots. Seed food reserves are stored outside the embryo, in the form of complex carbohydrates, lipids or proteins. The cotyledons serve as conduits to transmit the broken-down food reserves from their storage site inside the seed to the developing embryo. The seed consists of a toughened layer of integuments forming the coat, the endosperm with food reserves, and at the center, the well-protected embryo.
Most flowers are monoecious or bisexual, which means that they carry both stamens and carpels; only a few species self-pollinate. Monoecious flowers are also known as “perfect” flowers because they contain both types of sex organs (Figure 13). Both anatomical and environmental barriers promote cross-pollination mediated by a physical agent (wind or water), or an animal, such as an insect or bird. Cross-pollination increases genetic diversity in a species.
Diversity of Angiosperms
Angiosperms are classified in a single phylum: the Anthophyta. Modern angiosperms appear to be a monophyletic group, which means that they originate from a single ancestor. Flowering plants are divided into two major groups, according to the structure of the cotyledons, pollen grains, and other structures. Monocots include grasses and lilies, and eudicots or dicots form a polyphyletic group. Basal angiosperms are a group of plants that are believed to have branched off before the separation into monocots and eudicots because they exhibit traits from both groups. They are categorized separately in many classification schemes. The Magnoliidae (magnolia trees, laurels, and water lilies) and the Piperaceae (peppers) belong to the basal angiosperm group.
Basal Angiosperms
The Magnoliidae are represented by the magnolias: tall trees bearing large, fragrant flowers that have many parts and are considered archaic (Figure 15d). Laurel trees produce fragrant leaves and small, inconspicuous flowers. The Laurales grow mostly in warmer climates and are small trees and shrubs. Familiar plants in this group include the bay laurel, cinnamon, spice bush (Figure 15a), and avocado tree. The Nymphaeales are comprised of the water lilies, lotus (Figure 15c), and similar plants; all species thrive in freshwater biomes, and have leaves that float on the water surface or grow underwater. Water lilies are particularly prized by gardeners, and have graced ponds and pools for thousands of years. The Piperales are a group of herbs, shrubs, and small trees that grow in the tropical climates. They have small flowers without petals that are tightly arranged in long spikes. Many species are the source of prized fragrance or spices, for example the berries of Piper nigrum (Figure 4b) are the familiar black peppercorns that are used to flavor many dishes.

Figure 15. The (a) common spicebush belongs to the Laurales, the same family as cinnamon and bay laurel. The fruit of (b) the Piper nigrumplant is black pepper, the main product that was traded along spice routes. Notice the small, unobtrusive, clustered flowers. (c) Lotus flowers, Nelumbo nucifera, have been cultivated since ancient times for their ornamental value; the root of the lotus flower is eaten as a vegetable. The red seeds of (d) a magnolia tree, characteristic of the final stage, are just starting to appear. (credit a: modification of work by Cory Zanker; credit b: modification of work by Franz Eugen Köhler; credit c: modification of work by “berduchwal”/Flickr; credit d: modification of work by “Coastside2″/Wikimedia Commons).
Monocots
Plants in the monocot group are primarily identified as such by the presence of a single cotyledon in the seedling. Other anatomical features shared by monocots include veins that run parallel to the length of the leaves, and flower parts that are arranged in a three- or six-fold symmetry. True woody tissue is rarely found in monocots. In palm trees, vascular and parenchyma tissues produced by the primary and secondary thickening meristems form the trunk. The pollen from the first angiosperms was monosulcate, containing a single furrow or pore through the outer layer. This feature is still seen in the modern monocots. Vascular tissue of the stem is not arranged in any particular pattern. The root system is mostly adventitious and unusually positioned, with no major tap root. The monocots include familiar plants such as the true lilies (which are at the origin of their alternate name of Liliopsida), orchids, grasses, and palms. Many important crops are monocots, such as rice and other cereals, corn, sugar cane, and tropical fruits like bananas and pineapples (Figure16).

Figure 16. The world’s major crops are flowering plants. (a) Rice, (b) wheat, and (c) bananas are monocots, while (d) cabbage, (e) beans, and (f) peaches are dicots. (credit a: modification of work by David Nance, USDA ARS; credit b, c: modification of work by Rosendahl; credit d: modification of work by Bill Tarpenning, USDA; credit e: modification of work by Scott Bauer, USDA ARS; credit f: modification of work by Keith Weller, USDA)
Eudicots
Eudicots, or true dicots, are characterized by the presence of two cotyledons in the developing shoot. Veins form a network in leaves, and flower parts come in four, five, or many whorls. Vascular tissue forms a ring in the stem; in monocots, vascular tissue is scattered in the stem. Eudicots can be herbaceous (like grasses), or produce woody tissues. Most eudicots produce pollen that is trisulcate or triporate, with three furrows or pores. The root system is usually anchored by one main root developed from the embryonic radicle. Eudicots comprise two-thirds of all flowering plants. The major differences between monocots and eudicots are summarized in Table 1. Many species exhibit characteristics that belong to either group; as such, the classification of a plant as a monocot or a eudicot is not always clearly evident.
| Table 1. Comparison of Structural Characteristics of Monocots and Eudicots | ||
|---|---|---|
| Characteristic | Monocot | Eudicot |
| Cotyledon | One | Two |
| Veins in Leaves | Parallel | Network (branched) |
| Stem Vascular Tissue | Scattered | Arranged in ring pattern |
| Roots | Network of adventitious roots | Tap root with many lateral roots |
| Pollen | Monosulcate | Trisulcate |
| Flower Parts | Three or multiple of three | Four, five, multiple of four or five and whorls |
The Role of Seed Plants
Without seed plants, life as we know it would not be possible. Plants play a key role in the maintenance of terrestrial ecosystems through stabilization of soils, cycling of carbon, and climate moderation. Large tropical forests release oxygen and act as carbon dioxide sinks. Seed plants provide shelter to many life forms, as well as food for herbivores, thereby indirectly feeding carnivores. Plant secondary metabolites are used for medicinal purposes and industrial production.
Animals and Plants: Herbivory
Coevolution of flowering plants and insects is a hypothesis that has received much attention and support, especially because both angiosperms and insects diversified at about the same time in the middle Mesozoic. Many authors have attributed the diversity of plants and insects to pollination and herbivory, or consumption of plants by insects and other animals. This is believed to have been as much a driving force as pollination. Coevolution of herbivores and plant defenses is observed in nature. Unlike animals, most plants cannot outrun predators or use mimicry to hide from hungry animals. A sort of arms race exists between plants and herbivores. To “combat” herbivores, some plant seeds—such as acorn and unripened persimmon—are high in alkaloids and therefore unsavory to some animals. Other plants are protected by bark, although some animals developed specialized mouth pieces to tear and chew vegetal material. Spines and thorns (Figure 17) deter most animals, except for mammals with thick fur, and some birds have specialized beaks to get past such defenses.

Figure 17. (a) Spines and (b) thorns are examples of plant defenses. (credit a: modification of work by Jon Sullivan; credit b: modification of work by I. Sáček, Sr.)
Herbivory has been used by seed plants for their own benefit in a display of mutualistic relationships. The dispersal of fruit by animals is the most striking example. The plant offers to the herbivore a nutritious source of food in return for spreading the plant’s genetic material to a wider area.
An extreme example of collaboration between an animal and a plant is the case of acacia trees and ants. The trees support the insects with shelter and food. In return, ants discourage herbivores, both invertebrates and vertebrates, by stinging and attacking leaf-eating insects.
Animals and Plants: Pollination
Grasses are a successful group of flowering plants that are wind pollinated. They produce large amounts of powdery pollen carried over large distances by the wind. The flowers are small and wisp-like. Large trees such as oaks, maples, and birches are also wind pollinated.

Figure 18. As a bee collects nectar from a flower, it is dusted by pollen, which it then disperses to other flowers. (credit: John Severns)
More than 80 percent of angiosperms depend on animals for pollination: the transfer of pollen from the anther to the stigma. Consequently, plants have developed many adaptations to attract pollinators. The specificity of specialized plant structures that target animals can be very surprising. It is possible, for example, to determine the type of pollinator favored by a plant just from the flower’s characteristics. Many bird or insect-pollinated flowers secrete nectar, which is a sugary liquid.
They also produce both fertile pollen, for reproduction, and sterile pollen rich in nutrients for birds and insects. Butterflies and bees can detect ultraviolet light. Flowers that attract these pollinators usually display a pattern of low ultraviolet reflectance that helps them quickly locate the flower’s center and collect nectar while being dusted with pollen (Figure 18). Large, red flowers with little smell and a long funnel shape are preferred by hummingbirds, who have good color perception, a poor sense of smell, and need a strong perch. White flowers opened at night attract moths. Other animals—such as bats, lemurs, and lizards—can also act as pollinating agents. Any disruption to these interactions, such as the disappearance of bees as a consequence of colony collapse disorders, can lead to disaster for agricultural industries that depend heavily on pollinated crops.
Testing Attraction of Flies by Rotting Flesh Smell
Question: Will flowers that offer cues to bees attract carrion flies if sprayed with compounds that smell like rotten flesh?
Background: Visitation of flowers by pollinating flies is a function mostly of smell. Flies are attracted by rotting flesh and carrions. The putrid odor seems to be the major attractant. The polyamines putrescine and cadaverine, which are the products of protein breakdown after animal death, are the source of the pungent smell of decaying meat. Some plants strategically attract flies by synthesizing polyamines similar to those generated by decaying flesh and thereby attract carrion flies.
Flies seek out dead animals because they normally lay their eggs on them and their maggots feed on the decaying flesh. Interestingly, time of death can be determined by a forensic entomologist based on the stages and type of maggots recovered from cadavers.
Hypothesis: Because flies are drawn to other organisms based on smell and not sight, a flower that is normally attractive to bees because of its colors will attract flies if it is sprayed with polyamines similar to those generated by decaying flesh.
Test the hypothesis:
- Select flowers usually pollinated by bees. White petunia may be good choice.
- Divide the flowers into two groups, and while wearing eye protection and gloves, spray one group with a solution of either putrescine or cadaverine. (Putrescine dihydrochloride is typically available in 98 percent concentration; this can be diluted to approximately 50 percent for this experiment.)
- Place the flowers in a location where flies are present, keeping the sprayed and unsprayed flowers separated.
- Observe the movement of the flies for one hour. Record the number of visits to the flowers using a table similar to Table 1. Given the rapid movement of flies, it may be beneficial to use a video camera to record the fly–flower interaction. Replay the video in slow motion to obtain an accurate record of the number of fly visits to the flowers.
- Repeat the experiment four more times with the same species of flower, but using different specimens.
- Repeat the entire experiment with a different type of flower that is normally pollinated by bees.
| Table 1. Results of Number of Visits by Flies to Sprayed and Control/Unsprayed Flowers | ||
|---|---|---|
| Trial # | Sprayed Flowers | Unsprayed Flowers |
| 1 | ||
| 2 | ||
| 3 | ||
| 4 | ||
| 5 | ||
Analyze your data: Review the data you have recorded. Average the number of visits that flies made to sprayed flowers over the course of the five trials (on the first flower type) and compare and contrast them to the average number of visits that flies made to the unsprayed/control flowers. Can you draw any conclusions regarding the attraction of the flies to the sprayed flowers?
For the second flower type used, average the number of visits that flies made to sprayed flowers over the course of the five trials and compare and contrast them to the average number of visits that flies made to the unsprayed/control flowers. Can you draw any conclusions regarding the attraction of the flies to the sprayed flowers?
Compare and contrast the average number of visits that flies made to the two flower types. Can you draw any conclusions about whether the appearance of the flower had any impact on the attraction of flies? Did smell override any appearance differences, or were the flies attracted to one flower type more than another?
Form a conclusion: Do the results support the hypothesis? If not, how can this be explained?
The Importance of Seed Plants in Human Life
Seed plants are the foundation of human diets across the world (Figure 19). Many societies eat almost exclusively vegetarian fare and depend solely on seed plants for their nutritional needs. A few crops (rice, wheat, and potatoes) dominate the agricultural landscape. Many crops were developed during the agricultural revolution, when human societies made the transition from nomadic hunter–gatherers to horticulture and agriculture. Cereals, rich in carbohydrates, provide the staple of many human diets. Beans and nuts supply proteins. Fats are derived from crushed seeds, as is the case for peanut and rapeseed (canola) oils, or fruits such as olives. Animal husbandry also consumes large amounts of crops.
Staple crops are not the only food derived from seed plants. Fruits and vegetables provide nutrients, vitamins, and fiber. Sugar, to sweeten dishes, is produced from the monocot sugarcane and the eudicot sugar beet. Drinks are made from infusions of tea leaves, chamomile flowers, crushed coffee beans, or powdered cocoa beans. Spices come from many different plant parts: saffron and cloves are stamens and buds, black pepper and vanilla are seeds, the bark of a bush in the Laurales family supplies cinnamon, and the herbs that flavor many dishes come from dried leaves and fruit, such as the pungent red chili pepper. The volatile oils of flowers and bark provide the scent of perfumes. Additionally, no discussion of seed plant contribution to human diet would be complete without the mention of alcohol. Fermentation of plant-derived sugars and starches is used to produce alcoholic beverages in all societies. In some cases, the beverages are derived from the fermentation of sugars from fruit, as with wines and, in other cases, from the fermentation of carbohydrates derived from seeds, as with beers.
Seed plants have many other uses, including providing wood as a source of timber for construction, fuel, and material to build furniture. Most paper is derived from the pulp of coniferous trees. Fibers of seed plants such as cotton, flax, and hemp are woven into cloth. Textile dyes, such as indigo, were mostly of plant origin until the advent of synthetic chemical dyes.
Lastly, it is more difficult to quantify the benefits of ornamental seed plants. These grace private and public spaces, adding beauty and serenity to human lives and inspiring painters and poets alike.

Figure 19. Humans rely on plants for a variety of reasons. (a) Cacao beans were introduced in Europe from the New World, where they were used by Mesoamerican civilizations. Combined with sugar, another plant product, chocolate is a popular food. (b) Flowers like the tulip are cultivated for their beauty. (c) Quinine, extracted from cinchona trees, is used to treat malaria, to reduce fever, and to alleviate pain. (d) This violin is made of wood. (credit a: modification of work by “Everjean”/Flickr; credit b: modification of work by Rosendahl; credit c: modification of work by Franz Eugen Köhler)
The medicinal properties of plants have been known to human societies since ancient times. There are references to the use of plants’ curative properties in Egyptian, Babylonian, and Chinese writings from 5,000 years ago. Many modern synthetic therapeutic drugs are derived or synthesized de novo from plant secondary metabolites. It is important to note that the same plant extract can be a therapeutic remedy at low concentrations, become an addictive drug at higher doses, and can potentially kill at high concentrations. Table 2 presents a few drugs, their plants of origin, and their medicinal applications.
| Table 2. Plant Origin of Medicinal Compounds and Medical Applications | ||
|---|---|---|
| Plant | Compound | Application |
| Deadly nightshade (Atropa belladonna ) | Atropine | Dilate eye pupils for eye exams |
| Foxglove (Digitalis purpurea) | Digitalis | Heart disease, stimulates heart beat |
| Yam (Dioscorea spp.) | Steroids | Steroid hormones: contraceptive pill and cortisone |
| Ephedra (Ephedra spp.) | Ephedrine | Decongestant and bronchiole dilator |
| Pacific yew (Taxus brevifolia) | Taxol | Cancer chemotherapy; inhibits mitosis |
| Opium poppy (Papaver somniferum) | Opioids | Analgesic (reduces pain without loss of consciousness) and narcotic (reduces pain with drowsiness and loss of consciousness) in higher doses |
| Quinine tree (Cinchona spp.) | Quinine | Antipyretic (lowers body temperature) and antimalarial |
| Willow (Salix spp.) | Salicylic acid (aspirin) | Analgesic and antipyretic |
Biodiversity of Plants
Biodiversity ensures a resource for new food crops and medicines. Plant life balances ecosystems, protects watersheds, mitigates erosion, moderates climate and provides shelter for many animal species. Threats to plant diversity, however, come from many angles. The explosion of the human population, especially in tropical countries where birth rates are highest and economic development is in full swing, is leading to human encroachment into forested areas. To feed the larger population, humans need to obtain arable land, so there is massive clearing of trees. The need for more energy to power larger cities and economic growth therein leads to the construction of dams, the consequent flooding of ecosystems, and increased emissions of pollutants. Other threats to tropical forests come from poachers, who log trees for their precious wood. Ebony and Brazilian rosewood, both on the endangered list, are examples of tree species driven almost to extinction by indiscriminate logging.
The number of plant species becoming extinct is increasing at an alarming rate. Because ecosystems are in a delicate balance, and seed plants maintain close symbiotic relationships with animals—whether predators or pollinators—the disappearance of a single plant can lead to the extinction of connected animal species. A real and pressing issue is that many plant species have not yet been catalogued, and so their place in the ecosystem is unknown. These unknown species are threatened by logging, habitat destruction, and loss of pollinators. They may become extinct before we have the chance to begin to understand the possible impacts from their disappearance. Efforts to preserve biodiversity take several lines of action, from preserving heirloom seeds to barcoding species. Heirloom seeds come from plants that were traditionally grown in human populations, as opposed to the seeds used for large-scale agricultural production. Barcoding is a technique in which one or more short gene sequences, taken from a well-characterized portion of the genome, are used to identify a species through DNA analysis.
In Summary: Seed Plants
Seed plants appeared about one million years ago, during the Carboniferous period. Two major innovations—seed and pollen—allowed seed plants to reproduce in the absence of water. The gametophytes of seed plants shrank, while the sporophytes became prominent structures and the diploid stage became the longest phase of the lifecycle. Gymnosperms became the dominant group during the Triassic. In these, pollen grains and seeds protect against desiccation. The seed, unlike a spore, is a diploid embryo surrounded by storage tissue and protective layers. It is equipped to delay germination until growth conditions are optimal. Angiosperms bear both flowers and fruit. The structures protect the gametes and the embryo during its development. Angiosperms appeared during the Mesozoic era and have become the dominant plant life in terrestrial habitats.
Gymnosperms are heterosporous seed plants that produce naked seeds. They appeared in the Paleozoic period and were the dominant plant life during the Mesozoic. Modern-day gymnosperms belong to four phyla. The largest phylum, Coniferophyta, is represented by conifers, the predominant plants at high altitude and latitude. Cycads (phylum Cycadophyta) resemble palm trees and grow in tropical climates. Gingko biloba is the only representative of the phylum Gingkophyta. The last phylum, Gnetophyta, is a diverse group of shrubs that produce vessel elements in their wood.
Angiosperms are the dominant form of plant life in most terrestrial ecosystems, comprising about 90 percent of all plant species. Most crops and ornamental plants are angiosperms. Their success comes from two innovative structures that protect reproduction from variability in the environment: the flower and the fruit. Flowers were derived from modified leaves. The main parts of a flower are the sepals and petals, which protect the reproductive parts: the stamens and the carpels. The stamens produce the male gametes in pollen grains. The carpels contain the female gametes (the eggs inside the ovules), which are within the ovary of a carpel. The walls of the ovary thicken after fertilization, ripening into fruit that ensures dispersal by wind, water, or animals.
The angiosperm life cycle is dominated by the sporophyte stage. Double fertilization is an event unique to angiosperms. One sperm in the pollen fertilizes the egg, forming a diploid zygote, while the other combines with the two polar nuclei, forming a triploid cell that develops into a food storage tissue called the endosperm. Flowering plants are divided into two main groups, the monocots and eudicots, according to the number of cotyledons in the seedlings. Basal angiosperms belong to an older lineage than monocots and eudicots.
Angiosperm diversity is due in part to multiple interactions with animals. Herbivory has favored the development of defense mechanisms in plants, and avoidance of those defense mechanism in animals. Pollination (the transfer of pollen to a carpel) is mainly carried out by wind and animals, and angiosperms have evolved numerous adaptations to capture the wind or attract specific classes of animals.
Plants play a key role in ecosystems. They are a source of food and medicinal compounds, and provide raw materials for many industries. Rapid deforestation and industrialization, however, threaten plant biodiversity. In turn, this threatens the ecosystem.
Check Your Understanding
Answer the question(s) below to see how well you understand the topics covered in the previous section. This short quiz does not count toward your grade in the class, and you can retake it an unlimited number of times.
Use this quiz to check your understanding and decide whether to (1) study the previous section further or (2) move on to the next section.
Candela Citations
- Biology. Provided by: OpenStax CNX. Located at: http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8. License: CC BY: Attribution. License Terms: Download for free at http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8
- Introduction to Seed Plants. Authored by: Shelli Carter and Lumen Learning. Provided by: Lumen Learning. License: CC BY: Attribution
- "Geologic time scale," Wikipedia. https://en.wikipedia.org/wiki/Geologic_time_scale. ↵