Learning Outcomes
- Identify the role of the brain in muscle movement
Excitation–Contraction Coupling
Excitation–contraction coupling is the link (transduction) between the action potential generated in the sarcolemma and the start of a muscle contraction. The trigger for calcium release from the sarcoplasmic reticulum into the sarcoplasm is a neural signal. Each skeletal muscle fiber is controlled by a motor neuron, which conducts signals from the brain or spinal cord to the muscle. The area of the sarcolemma on the muscle fiber that interacts with the neuron is called the motor end plate. The end of the neuron’s axon is called the synaptic terminal, and it does not actually contact the motor end plate. A small space called the synaptic cleft separates the synaptic terminal from the motor end plate. Electrical signals travel along the neuron’s axon, which branches through the muscle and connects to individual muscle fibers at a neuromuscular junction.
The ability of cells to communicate electrically requires that the cells expend energy to create an electrical gradient across their cell membranes. This charge gradient is carried by ions, which are differentially distributed across the membrane. Each ion exerts an electrical influence and a concentration influence. Just as milk will eventually mix with coffee without the need to stir, ions also distribute themselves evenly, if they are permitted to do so. In this case, they are not permitted to return to an evenly mixed state.
The sodium–potassium ATPase uses cellular energy to move K+ ions inside the cell and Na+ ions outside. This alone accumulates a small electrical charge, but a big concentration gradient. There is lots of K+ in the cell and lots of Na+ outside the cell. Potassium is able to leave the cell through K+ channels that are open 90% of the time, and it does. However, Na+ channels are rarely open, so Na+ remains outside the cell. When K+ leaves the cell, obeying its concentration gradient, that effectively leaves a negative charge behind. So at rest, there is a large concentration gradient for Na+ to enter the cell, and there is an accumulation of negative charges left behind in the cell. This is the resting membrane potential. Potential in this context means a separation of electrical charge that is capable of doing work. It is measured in volts, just like a battery. However, the transmembrane potential is considerably smaller (0.07 V); therefore, the small value is expressed as millivolts (mV) or 70 mV. Because the inside of a cell is negative compared with the outside, a minus sign signifies the excess of negative charges inside the cell, −70 mV.
If an event changes the permeability of the membrane to Na+ ions, they will enter the cell. That will change the voltage. This is an electrical event, called an action potential, that can be used as a cellular signal. Communication occurs between nerves and muscles through neurotransmitters. Neuron action potentials cause the release of neurotransmitters from the synaptic terminal into the synaptic cleft, where they can then diffuse across the synaptic cleft and bind to a receptor molecule on the motor end plate. The motor end plate possesses junctional folds—folds in the sarcolemma that create a large surface area for the neurotransmitter to bind to receptors. The receptors are actually sodium channels that open to allow the passage of Na+ into the cell when they receive a neurotransmitter signal.
Acetylcholine (ACh) is a neurotransmitter released by motor neurons that binds to receptors in the motor end plate. Neurotransmitter release occurs when an action potential travels down the motor neuron’s axon, resulting in altered permeability of the synaptic terminal membrane and an influx of calcium. The Ca2+ ions allow synaptic vesicles to move to and bind with the presynaptic membrane (on the neuron), and release neurotransmitter from the vesicles into the synaptic cleft. Once released by the synaptic terminal, ACh diffuses across the synaptic cleft to the motor end plate, where it binds with ACh receptors. As a neurotransmitter binds, these ion channels open, and Na+ ions cross the membrane into the muscle cell. This reduces the voltage difference between the inside and outside of the cell, which is called depolarization. As ACh binds at the motor end plate, this depolarization is called an end-plate potential. The depolarization then spreads along the sarcolemma, creating an action potential as sodium channels adjacent to the initial depolarization site sense the change in voltage and open. The action potential moves across the entire cell, creating a wave of depolarization.
ACh is broken down by the enzyme acetylcholinesterase (AChE) into acetyl and choline. AChE resides in the synaptic cleft, breaking down ACh so that it does not remain bound to ACh receptors, which would cause unwanted extended muscle contraction (Figure 1).
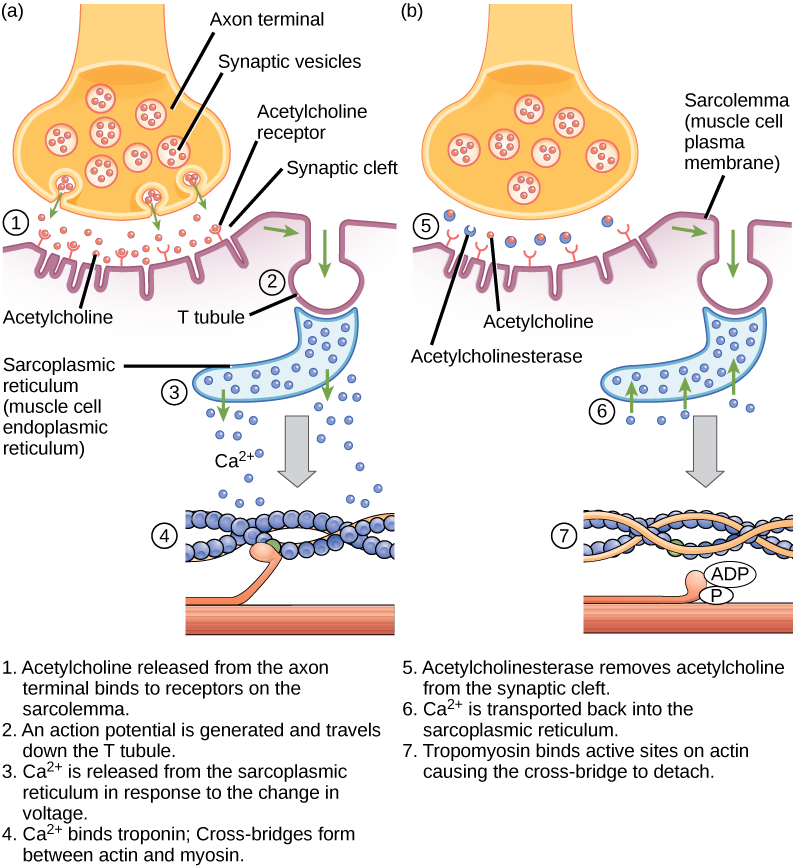
Figure 1. This diagram shows excitation-contraction coupling in a skeletal muscle contraction. The sarcoplasmic reticulum is a specialized endoplasmic reticulum found in muscle cells.
After depolarization, the membrane returns to its resting state. This is called repolarization, during which voltage-gated sodium channels close. Potassium channels continue at 90% conductance. Because the plasma membrane sodium–potassium ATPase always transports ions, the resting state (negatively charged inside relative to the outside) is restored. The period immediately following the transmission of an impulse in a nerve or muscle, in which a neuron or muscle cell regains its ability to transmit another impulse, is called the refractory period. During the refractory period, the membrane cannot generate another action potential. The refractory period allows the voltage-sensitive ion channels to return to their resting configurations. The sodium potassium ATPase continually moves Na+ back out of the cell and K+ back into the cell, and the K+ leaks out leaving negative charge behind. Very quickly, the membrane repolarizes, so that it can again be depolarized.
Practice Question
The deadly nerve gas Sarin irreversibly inhibits acetycholinesterase. What effect would Sarin have on muscle contraction?
Try It
Candela Citations
- Biology 2e. Provided by: OpenStax. Located at: http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8. License: CC BY: Attribution. License Terms: Access for free at https://openstax.org/books/biology-2e/pages/1-introduction
- Human Physiology. Provided by: Wikibooks. Located at: https://en.wikibooks.org/wiki/Human_Physiology. License: CC BY-SA: Attribution-ShareAlike
