Identify membrane-bound organelles found in eukaryotic cells
Have you ever heard the phrase “form follows function?” It’s a philosophy practiced in many industries. In architecture, this means that buildings should be constructed to support the activities that will be carried out inside them. For example, a skyscraper should be built with several elevator banks; a hospital should be built so that its emergency room is easily accessible.
Our natural world originated the principle of form following function, especially in cell biology, and this will become clear as we explore eukaryotic cells. Unlike prokaryotic cells, eukaryotic cells have: (1) a membrane-bound nucleus; (2) numerous membrane-bound organelles—such as the endoplasmic reticulum, Golgi apparatus, chloroplasts, mitochondria, and others; and (3) several, rod-shaped chromosomes. Because a eukaryotic cell’s nucleus is surrounded by a membrane, it is often said to have a “true nucleus.” The word “organelle” means “little organ,” and, as already mentioned, organelles have specialized cellular functions, just as the organs of your body have specialized functions.
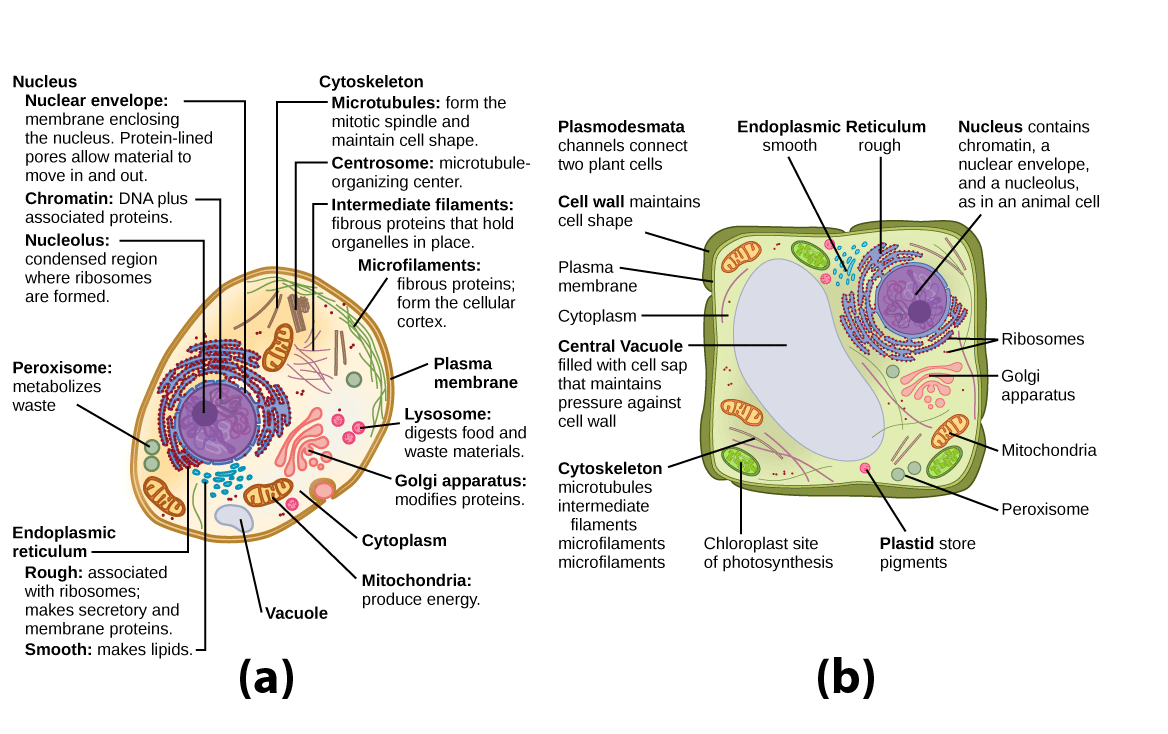
Figure 1. These figures show the major organelles and other cell components of (a) a typical animal cell and (b) a typical eukaryotic plant cell. The plant cell has a cell wall, chloroplasts, plastids, and a central vacuole—structures not found in animal cells. Plant cells do not have lysosomes or centrosomes.
Learning Objectives
- Describe the basic composition of cytoplasm
- Describe the structure and function of the nucleus and nuclear membrane
- Describe the structure and function of ribosomes
- Describe the structure and function of mitochondria
- Describe the structure and function of peroxisomes
- Describe the structure, function, and components of the endomembrane system
- Describe the structure and functions of vesicles
- Identify key organelles present only in animal cells, including centrosomes and lysosomes
- Identify key organelles present only in plant cells, including chloroplasts and vacuoles
Cytoplasm
Before we begin looking at individual organelles, we do need to briefly address the matrix in which they sit: the cytoplasm. The part of the cell referred to as cytoplasm is slightly different in eukaryotes and prokaryotes. In eukaryotic cells, which have a nucleus, the cytoplasm is everything between the plasma membrane and the nuclear envelope. In prokaryotes, which lack a nucleus, cytoplasm simply means everything found inside the plasma membrane.
One major component of the cytoplasm in both prokaryotes and eukaryotes is the gel-like cytosol, a water-based solution that contains ions, small molecules, and macromolecules. In eukaryotes, the cytoplasm also includes membrane-bound organelles, which are suspended in the cytosol. The cytoskeleton, a network of fibers that supports the cell and gives it shape, is also part of the cytoplasm and helps to organize cellular components.
Even though the cytosol is mostly water, it has a semi-solid, Jello-like consistency because of the many proteins suspended in it. The cytosol contains a rich broth of macromolecules and smaller organic molecules, including glucose and other simple sugars, polysaccharides, amino acids, nucleic acids, and fatty acids. Ions of sodium, potassium, calcium, and other elements are also found in the cytosol. Many metabolic reactions, including protein synthesis, take place in this part of the cell.
Nucleus

Figure 2. The nucleus stores chromatin (DNA plus proteins) in a gel-like substance called the nucleoplasm. The nucleolus is a condensed region of chromatin where ribosome synthesis occurs. The boundary of the nucleus is called the nuclear envelope. It consists of two phospholipid bilayers: an outer membrane and an inner membrane. The nuclear membrane is continuous with the endoplasmic reticulum. Nuclear pores allow substances to enter and exit the nucleus.
Typically, the nucleus is the most prominent organelle in a cell. The nucleus (plural = nuclei) houses the cell’s DNA and directs the synthesis of ribosomes and proteins. Let’s look at it in more detail (Figure 2).
The Nuclear Envelope
The nuclear envelope is a double-membrane structure that constitutes the outermost portion of the nucleus (Figure 2). Both the inner and outer membranes of the nuclear envelope are phospholipid bilayers.
The nuclear envelope is punctuated with pores that control the passage of ions, molecules, and RNA between the nucleoplasm and cytoplasm. The nucleoplasm is the semi-solid fluid inside the nucleus, where we find the chromatin and the nucleolus.
Chromatin and Chromosomes
To understand chromatin, it is helpful to first consider chromosomes. Chromosomes are structures within the nucleus that are made up of DNA, the hereditary material. In prokaryotes, DNA is organized into a single circular chromosome. In eukaryotes, chromosomes are linear structures. Every eukaryotic species has a specific number of chromosomes in the nuclei of its body’s cells. For example, in humans, the chromosome number is 46, while in fruit flies, it is eight. Chromosomes are only visible and distinguishable from one another when the cell is getting ready to divide. When the cell is in the growth and maintenance phases of its life cycle, proteins are attached to chromosomes, and they resemble an unwound, jumbled bunch of threads. These unwound protein-chromosome complexes are called chromatin (Figure 3); chromatin describes the material that makes up the chromosomes both when condensed and decondensed. We will focus on chromatin and chromosomes in greater detail later.

Figure 3. (a) This image shows various levels of the organization of chromatin (DNA and protein). (b) This image shows paired chromosomes. (credit b: modification of work by NIH; scale-bar data from Matt Russell)
The Nucleolus
We already know that the nucleus directs the synthesis of ribosomes, but how does it do this? Some chromosomes have sections of DNA that encode ribosomal RNA. A darkly staining area within the nucleus called the nucleolus (plural = nucleoli) aggregates the ribosomal RNA with associated proteins to assemble the ribosomal subunits that are then transported out through the pores in the nuclear envelope to the cytoplasm.
Ribosomes

Figure 4. Ribosomes are made up of a large subunit (top) and a small subunit (bottom). During protein synthesis, ribosomes assemble amino acids into proteins.
Ribosomes are the cellular structures responsible for protein synthesis. When viewed through an electron microscope, ribosomes appear either as clusters (polyribosomes) or single, tiny dots that float freely in the cytoplasm. They may be attached to the cytoplasmic side of the plasma membrane or the cytoplasmic side of the endoplasmic reticulum and the outer membrane of the nuclear envelope. Electron microscopy has shown us that ribosomes, which are large complexes of protein and RNA, consist of two subunits, aptly called large and small (Figure 4). Ribosomes receive their “orders” for protein synthesis from the nucleus where the DNA is transcribed into messenger RNA (mRNA). The mRNA travels to the ribosomes, which translate the code provided by the sequence of the nitrogenous bases in the mRNA into a specific order of amino acids in a protein. Amino acids are the building blocks of proteins.
Because proteins synthesis is an essential function of all cells, ribosomes are found in practically every cell. Ribosomes are particularly abundant in cells that synthesize large amounts of protein. For example, the pancreas is responsible for creating several digestive enzymes and the cells that produce these enzymes contain many ribosomes. Thus, we see another example of form following function.
Mitochondria
Mitochondria (singular = mitochondrion) are often called the “powerhouses” or “energy factories” of a cell because they are responsible for making adenosine triphosphate (ATP), the cell’s main energy-carrying molecule. ATP represents the short-term stored energy of the cell. Cellular respiration is the process of making ATP using the chemical energy found in glucose and other nutrients. In mitochondria, this process uses oxygen and produces carbon dioxide as a waste product. In fact, the carbon dioxide that you exhale with every breath comes from the cellular reactions that produce carbon dioxide as a byproduct.
In keeping with our theme of form following function, it is important to point out that muscle cells have a very high concentration of mitochondria that produce ATP. Your muscle cells need a lot of energy to keep your body moving. When your cells don’t get enough oxygen, they do not make a lot of ATP. Instead, the small amount of ATP they make in the absence of oxygen is accompanied by the production of lactic acid.

Figure 5. This electron micrograph shows a mitochondrion as viewed with a transmission electron microscope. This organelle has an outer membrane and an inner membrane. The inner membrane contains folds, called cristae, which increase its surface area. The space between the two membranes is called the intermembrane space, and the space inside the inner membrane is called the mitochondrial matrix. ATP synthesis takes place on the inner membrane. (credit: modification of work by Matthew Britton; scale-bar data from Matt Russell)
Mitochondria are oval-shaped, double membrane organelles (Figure 5) that have their own DNA and ribosomes (we’ll talk about these later!). Each membrane is a phospholipid bilayer embedded with proteins. The inner layer has folds called cristae. The area surrounded by the folds is called the mitochondrial matrix. The cristae and the matrix have different roles in cellular respiration.
Peroxisomes
Peroxisomes are small, round organelles enclosed by single membranes. They carry out oxidation reactions that break down fatty acids and amino acids. They also detoxify many poisons that may enter the body. Many of these oxidation reactions release hydrogen peroxide, H2O2, which would be damaging to cells; however, when these reactions are confined to peroxisomes, enzymes safely break down the H2O2 into oxygen and water. For example, alcohol is detoxified by peroxisomes in liver cells. Glyoxysomes, which are specialized peroxisomes in plants, are responsible for converting stored fats into sugars.
Endomembrane System

Figure 6. Membrane and secretory proteins are synthesized in the rough endoplasmic reticulum (RER). The RER’s membrane also sometimes modifies proteins. (credit: modification of work by Magnus Manske)
The endomembrane system (endo = “within”) is a group of membranes and organelles (Figure 6) in eukaryotic cells that works together to modify, package, and transport lipids and proteins. It includes the nuclear envelope, lysosomes, vesicles, and the endoplasmic reticulum and Golgi apparatus, which we will cover shortly. Although not technically within the cell, the plasma membrane is included in the endomembrane system because, as you will see, it interacts with the other endomembranous organelles. The endomembrane system does not include the membranes of either mitochondria or chloroplasts.
Figure 6 illustrates the connections of the endomembrane system as a (green) integral membrane protein in the ER is modified by attachment of a (purple) carbohydrate. Vesicles with the integral protein bud from the ER and fuse with the cis face of the Golgi apparatus. As the protein passes along the Golgi’s cisternae, it is further modified by the addition of more carbohydrates. After its synthesis is complete, it exits as integral membrane protein of the vesicle that bud from the Golgi’s trans face and when the vesicle fuses with the cell membrane the protein becomes integral portion of that cell membrane.
The Endoplasmic Reticulum
The endoplasmic reticulum (ER) (Figure 6) is a series of interconnected membranous sacs and tubules that collectively modifies proteins and synthesizes lipids. However, these two functions are performed in separate areas of the ER: the rough ER and the smooth ER, respectively.
The hollow portion of the ER tubules is called the lumen or cisternal space. The membrane of the ER, which is a phospholipid bilayer embedded with proteins, is continuous with the nuclear envelope.
Rough ER
The rough endoplasmic reticulum (RER) is so named because the ribosomes attached to its cytoplasmic surface give it a studded appearance when viewed through an electron microscope (Figure 7).

Figure 7. This transmission electron micrograph shows the rough endoplasmic reticulum and other organelles in a pancreatic cell. (credit: modification of work by Louisa Howard)
Ribosomes transfer their newly synthesized proteins into the lumen of the RER where they undergo structural modifications, such as folding or the acquisition of side chains. These modified proteins will be incorporated into cellular membranes—the membrane of the ER or those of other organelles—or secreted from the cell (such as protein hormones or enzymes). The RER also makes phospholipids for cellular membranes.
If the phospholipids or modified proteins are not destined to stay in the RER, they will reach their destinations via transport vesicles that bud from the RER’s membrane (Figure 6).
Since the RER is engaged in modifying proteins (such as enzymes, for example) that will be secreted from the cell, you would be correct in assuming that the RER is abundant in cells that secrete proteins. This is the case with cells of the liver, for example.
Smooth ER
The smooth endoplasmic reticulum (SER) is continuous with the RER but has few or no ribosomes on its cytoplasmic surface. Functions of the SER include synthesis of carbohydrates, lipids, and steroid hormones; detoxification of medications and poisons; and storage of calcium ions.
In muscle cells, a specialized SER called the sarcoplasmic reticulum is responsible for storage of the calcium ions that are needed to trigger the coordinated contractions of the muscle cells.
Cardiology
Heart disease is the leading cause of death in the United States. This is primarily due to our sedentary lifestyle and our high trans-fat diets.
Heart failure is just one of many disabling heart conditions. Heart failure does not mean that the heart has stopped working. Rather, it means that the heart can’t pump with sufficient force to transport oxygenated blood to all the vital organs. Left untreated, heart failure can lead to kidney failure and failure of other organs.
The wall of the heart is composed of cardiac muscle tissue. Heart failure occurs when the endoplasmic reticula of cardiac muscle cells do not function properly. As a result, an insufficient number of calcium ions are available to trigger a sufficient contractile force.
Cardiologists (cardi = “heart”; ologist = “one who studies”) are doctors who specialize in treating heart diseases, including heart failure. Cardiologists can make a diagnosis of heart failure via physical examination, results from an electrocardiogram (ECG, a test that measures the electrical activity of the heart), a chest X-ray to see whether the heart is enlarged, and other tests. If heart failure is diagnosed, the cardiologist will typically prescribe appropriate medications and recommend a reduction in table salt intake and a supervised exercise program.
The Golgi Apparatus

Figure 8. The Golgi apparatus in this white blood cell is visible as a stack of semicircular, flattened rings in the lower portion of the image. Several vesicles can be seen near the Golgi apparatus. (credit: modification of work by Louisa Howard)
We have already mentioned that vesicles can bud from the ER and transport their contents elsewhere, but where do the vesicles go? Before reaching their final destination, the lipids or proteins within the transport vesicles still need to be sorted, packaged, and tagged so that they wind up in the right place. Sorting, tagging, packaging, and distribution of lipids and proteins takes place in the Golgi apparatus (also called the Golgi body), a series of flattened membranes (Figure 8).
The receiving side of the Golgi apparatus is called the cis face. The opposite side is called the trans face. The transport vesicles that formed from the ER travel to the cis face, fuse with it, and empty their contents into the lumen of the Golgi apparatus. As the proteins and lipids travel through the Golgi, they undergo further modifications that allow them to be sorted. The most frequent modification is the addition of short chains of sugar molecules. These newly modified proteins and lipids are then tagged with phosphate groups or other small molecules so that they can be routed to their proper destinations.
Finally, the modified and tagged proteins are packaged into secretory vesicles that bud from the trans face of the Golgi. While some of these vesicles deposit their contents into other parts of the cell where they will be used, other secretory vesicles fuse with the plasma membrane and release their contents outside the cell.
In another example of form following function, cells that engage in a great deal of secretory activity (such as cells of the salivary glands that secrete digestive enzymes or cells of the immune system that secrete antibodies) have an abundance of Golgi.
In plant cells, the Golgi apparatus has the additional role of synthesizing polysaccharides, some of which are incorporated into the cell wall and some of which are used in other parts of the cell.
Geneticist
Many diseases arise from genetic mutations that prevent the synthesis of critical proteins. One such disease is Lowe disease (also called oculocerebrorenal syndrome, because it affects the eyes, brain, and kidneys). In Lowe disease, there is a deficiency in an enzyme localized to the Golgi apparatus. Children with Lowe disease are born with cataracts, typically develop kidney disease after the first year of life, and may have impaired mental abilities.
Lowe disease is a genetic disease caused by a mutation on the X chromosome. The X chromosome is one of the two human sex chromosome, as these chromosomes determine a person’s sex. Females possess two X chromosomes while males possess one X and one Y chromosome. In females, the genes on only one of the two X chromosomes are expressed. Therefore, females who carry the Lowe disease gene on one of their X chromosomes have a 50/50 chance of having the disease. However, males only have one X chromosome and the genes on this chromosome are always expressed. Therefore, males will always have Lowe disease if their X chromosome carries the Lowe disease gene. The location of the mutated gene, as well as the locations of many other mutations that cause genetic diseases, has now been identified. Through prenatal testing, a woman can find out if the fetus she is carrying may be afflicted with one of several genetic diseases.
Geneticists analyze the results of prenatal genetic tests and may counsel pregnant women on available options. They may also conduct genetic research that leads to new drugs or foods, or perform DNA analyses that are used in forensic investigations.
Vesicles
Vesicles, like vacuoles (which we will address soon), are membrane-bound sacs that function in storage and transport. Other than the fact that vacuoles are somewhat larger than vesicles, there is a very subtle distinction between them: the membranes of vesicles can fuse with either the plasma membrane or other membrane systems within the cell. The membrane of a vacuole does not fuse with the membranes of other cellular components.
A vesicle consists of fluid enclosed by a lipid bilayer. Vesicles form naturally during the processes of secretion (exocytosis), uptake (phagocytosis) and transport of materials within the cytoplasm. Alternatively, they may be prepared artificially, in which case they are called liposomes. Vesicles can fuse with the plasma membrane to release their contents outside the cell. Vesicles can also fuse with other organelles within the cell.
Vesicles perform a variety of functions. Because they are separated from the cytosol, the inside of a vesicle can be different from the cytosolic environment. For this reason, vesicles are a basic tool used by the cell for organizing cellular substances. Vesicles are involved in metabolism, transport, buoyancy control, and enzyme storage. They can also act as chemical reaction chambers.
Unique Features of Animal and Plant Cells
At this point, you know that each eukaryotic cell has a plasma membrane, cytoplasm, a nucleus, ribosomes, mitochondria, peroxisomes, and in some, vacuoles, but there are some striking differences between animal and plant cells. While both animal and plant cells have microtubule organizing centers (MTOCs), animal cells also have centrioles associated with the MTOC: a complex called the centrosome. Animal cells each have a centrosome and lysosomes, whereas plant cells do not. Plant cells have a cell wall, chloroplasts and other specialized plastids, and a large central vacuole, whereas animal cells do not.
Properties of Animal Cells

Figure 9. The centrosome consists of two centrioles that lie at right angles to each other. Each centriole is a cylinder made up of nine triplets of microtubules. Nontubulin proteins (indicated by the green lines) hold the microtubule triplets together.
The Centrosome
The centrosome is a microtubule-organizing center found near the nuclei of animal cells. It contains a pair of centrioles, two structures that lie perpendicular to each other (Figure 9). Each centriole is a cylinder of nine triplets of microtubules.
The centrosome (the organelle where all microtubules originate) replicates itself before a cell divides, and the centrioles appear to have some role in pulling the duplicated chromosomes to opposite ends of the dividing cell. However, the exact function of the centrioles in cell division isn’t clear, because cells that have had the centrosome removed can still divide, and plant cells, which lack centrosomes, are capable of cell division.
Lysosomes

Figure 10. A macrophage has engulfed (phagocytized) a potentially pathogenic bacterium and then fuses with a lysosomes within the cell to destroy the pathogen. Other organelles are present in the cell but for simplicity are not shown.
In addition to their role as the digestive component and organelle-recycling facility of animal cells, lysosomes are considered to be parts of the endomembrane system.
Lysosomes also use their hydrolytic enzymes to destroy pathogens (disease-causing organisms) that might enter the cell. A good example of this occurs in a group of white blood cells called macrophages, which are part of your body’s immune system. In a process known as phagocytosis or endocytosis, a section of the plasma membrane of the macrophage invaginates (folds in) and engulfs a pathogen. The invaginated section, with the pathogen inside, then pinches itself off from the plasma membrane and becomes a vesicle. The vesicle fuses with a lysosome. The lysosome’s hydrolytic enzymes then destroy the pathogen (Figure 10).
Properties of Plant Cells
Chloroplasts
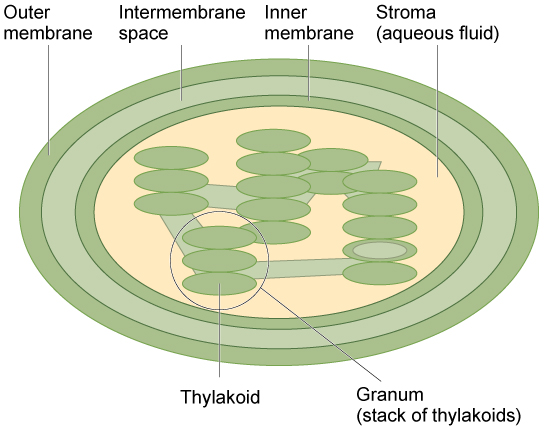
Figure 11. The chloroplast has an outer membrane, an inner membrane, and membrane structures called thylakoids that are stacked into grana. The space inside the thylakoid membranes is called the thylakoid space. The light harvesting reactions take place in the thylakoid membranes, and the synthesis of sugar takes place in the fluid inside the inner membrane, which is called the stroma. Chloroplasts also have their own genome, which is contained on a single circular chromosome.
Like the mitochondria, chloroplasts have their own DNA and ribosomes (we’ll talk about these later!), but chloroplasts have an entirely different function. Chloroplasts are plant cell organelles that carry out photosynthesis. Photosynthesis is the series of reactions that use carbon dioxide, water, and light energy to make glucose and oxygen. This is a major difference between plants and animals; plants (autotrophs) are able to make their own food, like sugars, while animals (heterotrophs) must ingest their food.
Like mitochondria, chloroplasts have outer and inner membranes, but within the space enclosed by a chloroplast’s inner membrane is a set of interconnected and stacked fluid-filled membrane sacs called thylakoids (Figure 11). Each stack of thylakoids is called a granum (plural = grana). The fluid enclosed by the inner membrane that surrounds the grana is called the stroma.
The chloroplasts contain a green pigment called chlorophyll, which captures the light energy that drives the reactions of photosynthesis. Like plant cells, photosynthetic protists also have chloroplasts. Some bacteria perform photosynthesis, but their chlorophyll is not relegated to an organelle.
Endosymbiosis
We have mentioned that both mitochondria and chloroplasts contain DNA and ribosomes. Have you wondered why? Strong evidence points to endosymbiosis as the explanation.
Symbiosis is a relationship in which organisms from two separate species depend on each other for their survival. Endosymbiosis (endo– = “within”) is a mutually beneficial relationship in which one organism lives inside the other. Endosymbiotic relationships abound in nature. We have already mentioned that microbes that produce vitamin K live inside the human gut. This relationship is beneficial for us because we are unable to synthesize vitamin K. It is also beneficial for the microbes because they are protected from other organisms and from drying out, and they receive abundant food from the environment of the large intestine.
Scientists have long noticed that bacteria, mitochondria, and chloroplasts are similar in size. We also know that bacteria have DNA and ribosomes, just as mitochondria and chloroplasts do. Scientists believe that host cells and bacteria formed an endosymbiotic relationship when the host cells ingested both aerobic and autotrophic bacteria (cyanobacteria) but did not destroy them. Through many millions of years of evolution, these ingested bacteria became more specialized in their functions, with the aerobic bacteria becoming mitochondria and the autotrophic bacteria becoming chloroplasts.
Vacuoles
Vacuoles are membrane-bound sacs that function in storage and transport. The membrane of a vacuole does not fuse with the membranes of other cellular components. Additionally, some agents such as enzymes within plant vacuoles break down macromolecules.
The Central Vacuole
If you look at Figure 1b, you will see that plant cells each have a large central vacuole that occupies most of the area of the cell. The central vacuole plays a key role in regulating the cell’s concentration of water in changing environmental conditions. Have you ever noticed that if you forget to water a plant for a few days, it wilts? That’s because as the water concentration in the soil becomes lower than the water concentration in the plant, water moves out of the central vacuoles and cytoplasm. As the central vacuole shrinks, it leaves the cell wall unsupported. This loss of support to the cell walls of plant cells results in the wilted appearance of the plant.
The central vacuole also supports the expansion of the cell. When the central vacuole holds more water, the cell gets larger without having to invest a lot of energy in synthesizing new cytoplasm. You can rescue wilted celery in your refrigerator using this process. Simply cut the end off the stalks and place them in a cup of water. Soon the celery will be stiff and crunchy again.
Check Your Understanding
Answer the question(s) below to see how well you understand the topics covered in the previous section. This short quiz does not count toward your grade in the class, and you can retake it an unlimited number of times.
Use this quiz to check your understanding and decide whether to (1) study the previous section further or (2) move on to the next section.
Candela Citations
- Revision, adaptation, and original content. Authored by: Shelli Carter and Lumen Learning. Provided by: Lumen Learning. License: CC BY: Attribution
- Biology. Provided by: OpenStax CNX. Located at: http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8. License: CC BY: Attribution. License Terms: Download for free at http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8
- Plasma membrane and cytoplasm. Provided by: Khan Academy. Located at: https://www.khanacademy.org/science/biology/structure-of-a-cell/prokaryotic-and-eukaryotic-cells/a/plasma-membrane-and-cytoplasm. Project: Biology. License: CC BY-NC-SA: Attribution-NonCommercial-ShareAlike
- Vesicles and Vacuoles. Provided by: Boundless. Located at: https://www.boundless.com/biology/textbooks/boundless-biology-textbook/cell-structure-4/the-endomembrane-system-and-proteins-61/vesicles-and-vacuoles-1004-17181/. License: CC BY-SA: Attribution-ShareAlike