Learning Outcomes
- Understand past and emerging classification systems for viruses
Because most viruses probably evolved from different ancestors, the systematic methods that scientists have used to classify prokaryotic and eukaryotic cells are not very useful. If viruses represent “remnants” of different organisms, then even genomic or protein analysis is not useful. Why?, Because viruses have no common genomic sequence that they all share. For example, the 16S rRNA sequence so useful for constructing prokaryote phylogenies is of no use for a creature with no ribosomes! Biologists have used several classification systems in the past. Viruses were initially grouped by shared morphology. Later, groups of viruses were classified by the type of nucleic acid they contained, DNA or RNA, and whether their nucleic acid was single- or double-stranded. However, these earlier classification methods grouped viruses differently, because they were based on different sets of characters of the virus. The most commonly used classification method today is called the Baltimore classification scheme, and is based on how messenger RNA (mRNA) is generated in each particular type of virus.
Past Systems of Classification
Viruses contain only a few elements by which they can be classified: the viral genome, the type of capsid, and the envelope structure for the enveloped viruses. All of these elements have been used in the past for viral classification (Table 1 and Figure 1). Viral genomes may vary in the type of genetic material (DNA or RNA) and its organization (single- or double-stranded, linear or circular, and segmented or non-segmented). In some viruses, additional proteins needed for replication are associated directly with the genome or contained within the viral capsid.
| Table 1. Virus Classification by Genome Structure and Core | |
|---|---|
| Core Classifications | Examples |
| RNA | Rabies virus, retroviruses |
| DNA | Herpesviruses, smallpox virus |
| Single-stranded | Rabies virus, retroviruses |
| Double-stranded | Herpesviruses, smallpox virus |
| Linear | Rabies virus, retroviruses, herpesviruses, smallpox virus |
| Circular | Papillomaviruses, many bacteriophages |
| Non-segmented: genome consists of a single segment of genetic material | Parainfluenza viruses |
| Segmented: genome is divided into multiple segments | Influenza viruses |
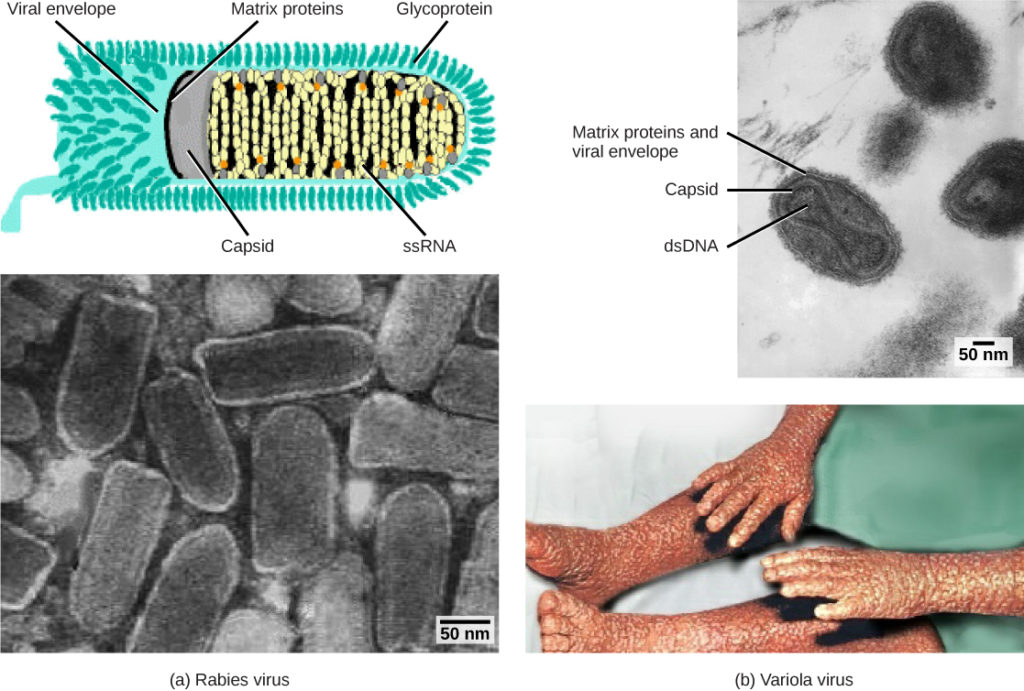
Figure 1. Viruses are classified based on their core genetic material and capsid design. (a) Rabies virus has a single-stranded RNA (ssRNA) core and an enveloped helical capsid, whereas (b) variola virus, the causative agent of smallpox, has a double-stranded DNA (dsDNA) core and a complex capsid. (credit “rabies diagram”: modification of work by CDC; “rabies micrograph”: modification of work by Dr. Fred Murphy, CDC; credit “small pox micrograph”: modification of work by Dr. Fred Murphy, Sylvia Whitfield, CDC; credit “smallpox photo”: modification of work by CDC; scale-bar data from Matt Russell)
Viruses can also be classified by the design of their capsids (Table 2 and Figure 2). Capsids are classified as naked icosahedral, enveloped icosahedral, enveloped helical, naked helical, and complex. The type of genetic material (DNA or RNA) and its structure (single- or double-stranded, linear or circular, and segmented or non-segmented) are used to classify the virus core structures (Table 2).
| Table 2. Virus Classification by Capsid Structure | |
|---|---|
| Capsid Classification | Examples |
| Naked icosahedral | Hepatitis A virus, polioviruses |
| Enveloped icosahedral | Epstein-Barr virus, herpes simplex virus, rubella virus, yellow fever virus, HIV-1 |
| Enveloped helical | Influenza viruses, mumps virus, measles virus, rabies virus |
| Naked helical | Tobacco mosaic virus |
| Complex with many proteins; some have combinations of icosahedral and helical capsid structures | Herpesviruses, smallpox virus, hepatitis B virus, T4 bacteriophage |

Figure 2. Transmission electron micrographs of various viruses show their structures. The capsid of the (a) polio virus is naked icosahedral; (b) the Epstein-Barr virus capsid is enveloped icosahedral; (c) the mumps virus capsid is an enveloped helix; (d) the tobacco mosaic virus capsid is naked helical; and (e) the herpesvirus capsid is complex. (credit a: modification of work by Dr. Fred Murphy, Sylvia Whitfield; credit b: modification of work by Liza Gross; credit c: modification of work by Dr. F. A. Murphy, CDC; credit d: modification of work by USDA ARS; credit e: modification of work by Linda Stannard, Department of Medical Microbiology, University of Cape Town, South Africa, NASA; scale-bar data from Matt Russell)
Baltimore Classification
The most commonly used system of virus classification was developed by Nobel Prize-winning biologist David Baltimore in the early 1970s. In addition to the differences in morphology and genetics mentioned above, the Baltimore classification scheme groups viruses according to how the mRNA is produced during the replicative cycle of the virus.
Group I viruses contain double-stranded DNA (dsDNA) as their genome. Their mRNA is produced by transcription in much the same way as with cellular DNA.
Group II viruses have single-stranded DNA (ssDNA) as their genome. They convert their single-stranded genomes into a dsDNA intermediate before transcription to mRNA can occur.
Group III viruses use dsRNA as their genome. The strands separate, and one of them is used as a template for the generation of mRNA using the RNA-dependent RNA polymerase encoded by the virus.
Group IV viruses have ssRNA as their genome with a positive polarity. Positive polarity means that the genomic RNA can serve directly as mRNA. Intermediates of dsRNA, called replicative intermediates, are made in the process of copying the genomic RNA. Multiple, full-length RNA strands of negative polarity (complimentary to the positive-stranded genomic RNA) are formed from these intermediates, which may then serve as templates for the production of RNA with positive polarity, including both full-length genomic RNA and shorter viral mRNAs.
Group V viruses contain ssRNA genomes with a negative polarity, meaning that their sequence is complementary to the mRNA. As with Group IV viruses, dsRNA intermediates are used to make copies of the genome and produce mRNA. In this case, the negative-stranded genome can be converted directly to mRNA. Additionally, full-length positive RNA strands are made to serve as templates for the production of the negative-stranded genome.
Group VI viruses have diploid (two copies) ssRNA genomes that must be converted, using the enzyme reverse transcriptase, to dsDNA; the dsDNA is then transported to the nucleus of the host cell and inserted into the host genome. Then, mRNA can be produced by transcription of the viral DNA that was integrated into the host genome.
Group VII viruses have partial dsDNA genomes and make ssRNA intermediates that act as mRNA, but are also converted back into dsDNA genomes by reverse transcriptase, necessary for genome replication. The characteristics of each group in the Baltimore classification are summarized in Table 3 with examples of each group.
| Table 3. Baltimore Classification | |||
|---|---|---|---|
| Group | Characteristics | Mode of mRNA Production | Example |
| I | Double-stranded DNA | mRNA is transcribed directly from the DNA template | Herpes simplex (herpesvirus) |
| II | Single-stranded DNA | DNA is converted to double-stranded form before RNA is transcribed | Canine parvovirus (parvovirus) |
| III | Double-stranded RNA | mRNA is transcribed from the RNA genome | Childhood gastroenteritis (rotavirus) |
| IV | Single stranded RNA (+) | Genome functions as mRNA | Common cold (pircornavirus) |
| V | Single stranded RNA (−) | mRNA is transcribed from the RNA genome | Rabies (rhabdovirus) |
| VI | Single stranded RNA viruses with reverse transcriptase | Reverse transcriptase makes DNA from the RNA genome; DNA is then incorporated in the host genome; mRNA is transcribed from the incorporated DNA | Human immunodeficiency virus (HIV) |
| VII | Double stranded DNA viruses with reverse transcriptase | The viral genome is double-stranded DNA, but viral DNA is replicated through an RNA intermediate; the RNA may serve directly as mRNA or as a template to make mRNA | Hepatitis B virus (hepadnavirus) |
Try It
Candela Citations
- Biology 2e. Provided by: OpenStax. Located at: http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8. License: CC BY: Attribution. License Terms: Access for free at https://openstax.org/books/biology-2e/pages/1-introduction
